3.1. Scope for bioethanol production from macroalgal biomass
The availability of spectrum of species amenable for biofuel production is one of the unique aspect of algae when compared to other advanced feedstock [35]. Production of different types of biofuels (bioethanol, diesel, bio-oil, and bio-hydrogen) is optimized by prioritizing species.
Characteristics of prioritized macroalgal feedstock for sustainable biofuel production are
(i) availability throughout the year (during all seasons)
(ii) presence of higher quantum of sugar.
3.1.1. Macroalgal feedstock availability and selection
Indian coast has about 1153 marine algal species belonging to 271 genera, of which 60 species are commercially important. Rocky beaches, mudflats, estuaries, coral reefs and lagoons are the preferred habitats for macroalgae. Indian coast harbors predominantly intertidal and subtidal algal communities [143]. Macroalgal feedstock (for bioethanol production) distribution along the Indian Coast and islands were compiled from various secondary data sources [144–150] along with the locations based on the current field investigations is represented in Fig. 4. Seaweed species belonging to Laminaria genera (Brown algae or Phaeophyta) are not recorded in India and these are algae of cold waters and certain brown algae of orders Dictyotales and Fucales (e.g. Sargassum) are distinctly warm-water plants [151] are abundantly spread and are recorded from Indian Coast. Kappaphycus alvarezii, a red algae is native of Indonesia and Philippines and introduced in India for commercial cultivation purpose [152], which has now colonized in various parts of Gulf of Mannar.
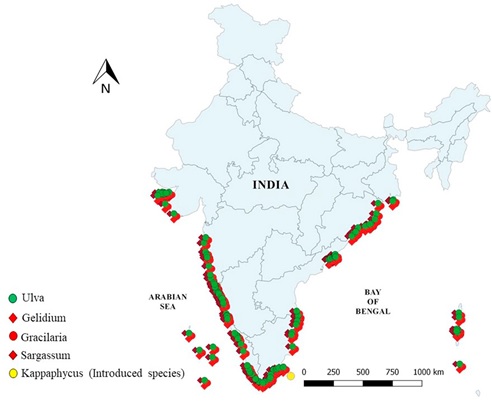 Fig. 4. Distribution of potential macroalgal feedstock of Indian coast.
Fig. 4. Distribution of potential macroalgal feedstock of Indian coast.
Setting up of bio refinery necessitates easier cultivation strategies apart from ensuring the availability of feedstock during all seasons. During the current field investigations covering 24 months, 25 seaweed species belonging to 19 genera were recorded from intertidal zone.
Among these, eight seaweed species abundantly grows during post- monsoon season in the Central West Coast. These species include Enteromorpha intestinalis (938.5 g/m
2), Ulva lactuca (1024.5 g/m
2), Chaetomorpha media (441.8 g/m
2), Gracilaria corticata (1039.5 g/m
2), Gelidium pusillum (205.4 g/m
2), Grateloupia lithophila (196.6 g/m
2), Sargassum ilcifolium (1175.7 g/m
2) and Padina tetrastromatica (1506.7 g/ m
2), which were considered for further analysis of biochemical composition (Fig. 5). It was seen that green seaweeds; U.lactuca and E. intestinalis, and red seaweed G.corticata occurred in sufficient quantity during all seasons. Occurrence of Ulva and Enteromorpha in all the seasons is attributed to their euryhaline nature, whereas G.corticata species are strictly marine and are restricted to open ocean environment [153]. Year round optimal biomass production of these macroalgal species, overcomes the seasonal constraints faced by the first and second generation biofuel feedstock [45].
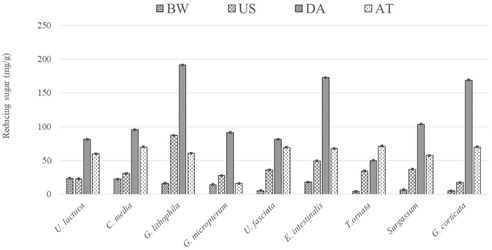 Fig. 5. Reducing sugar (mgg -1) extracted using various pretreatment methods for selected seaweeds.
Fig. 5. Reducing sugar (mgg -1) extracted using various pretreatment methods for selected seaweeds.
3.1.2. Biochemical composition of selected seaweeds
Macroalgal species with the higher quantum of sugar plays an important role as the composition of sugar influences the ethanol yield. Biochemical composition of chosen macroalgae were carried out focusing mainly on higher carbohydrate content which is an essential parameter for prioritizing feedstock for bioethanol production (Table 5). Algal biomass are composed of large quantities of carbohydrates in the cell wall mostly in the form of structural (cellulose) and storage (starch) polysaccharides, that serve as substrate for fermentation. Highest total carbohydrate was recorded from green seaweed Ulva lactuca (62.15 12.8%) followed by G. lithophila (60.5 11.4%), G.pusilum (50.1 3.4%) and E.intestinalis (40.1 14.6%). Among these, G.lithophila and G.pusilum have lower biomass production as compared to Ulva and Enteromorpha, which makes them unsuitable as feedstock for biofuel production. Protein ranged from 3.7 0.94 to 27.3 15.21%. Highest protein content was recorded from C. media (27.3 15.21%) followed by E. intestinalis (20.4 0.67%) and U. lactuca (17.3 1.68%). In green seaweed Ulva, protein concentration ranged from 9 to 33%. Lowest protein recorded was from G. lithophila (3.7 0.94%). In general, protein fraction is lower in brown seaweeds (3–15% DW) compared to green or red seaweeds (10–47% DW) similar to the earlier report [154]. Lipid ranged from 0.5 0.2 to 6 4.4%. Highest lipid was estimated in U. lactuca (6 4.4%) followed by G. pusillum (3.7 1.4%), P. tetrastromatica (2.8 0.3%). Lowest lipid content estimated in G. corticata (0.5 0.2%). Transesterification of lipids extracted from Enteromorpha biomass yielded 90.6% biodiesel [135], which highlights that Enteromorpha as a promising feedstock for biodiesel production.
| Seaweeds |
Total carbohydrate |
Soluble carbohydrate |
Insoluble carbohydrate |
Protein |
Lipid |
Cellulose |
C |
H |
N |
| U.lactuca |
62.15± 12.8 |
5.5± 0.07 |
56.5± 12.7 |
17.3± 1.68 |
6± 4.4 |
14.03± 0.14 |
25.31 |
5.44 |
2.61 |
| E.intestinalis |
40.1± 14.6 |
7.5± 0.02 |
32.5± 14.5 |
20.4± 0.67 |
2.8± 0.1 |
12.10± 0.53 |
33.00 |
6.44 |
4.36 |
| C.media |
25.5± 14.6 |
5.3± 0.02 |
20.1± 11 |
27.3±15.21 |
0.6± 0.01 |
10.53± 0.17 |
30.14 |
5.31 |
3.28 |
| G.corticata |
28.2± 11.1 |
13.4± 0.01 |
14.7± 11 |
14.4±1.33 |
0.5± 0.2 |
0.87± 0.07 |
26.46 |
5.01 |
1.89 |
| G. pusillum |
50.1±3.4 |
4.7± 0.01 |
45.4± 3.4 |
8±2.04 |
3.7± 1.4 |
1.55± 0.05 |
37.95 |
6.48 |
3.45 |
| G. lithophila |
60.5±11.4 |
14.8± 0.01 |
45.7± 11 |
3.7±0.94 |
2.4± 1.3 |
6.23± 0.16 |
29.60 |
6.15 |
2.52 |
| S. ilcifolium |
26.4±13.4 |
3.6± 0.01 |
22.7± 13 |
12.4±0.67 |
1.9± 0.01 |
1.30± 0.09 |
26.20 |
4.25 |
1.88 |
| P.tetrastromatica |
32.5±13.23 |
3.5± 0.02 |
28.9± 13.2 |
9.7±1.76 |
2.8± 0.3 |
1.48± 0.07 |
30.68 |
5.54 |
2.35 |
Table 5 Biochemical composition (%) of seaweeds along Central West Coast of India.
Elemental analysis of the seaweeds indicates 25.31–37.95% of carbon, 4.52–6.48% hydrogen and 1.88–4.36% Nitrogen. Highest carbon, hydrogen and nitrogen content were recorded respectively from G. pusillum (C: 37.95%), G.pusillum (H: 6.48%) and E.intestinalis (N: 4.36%) respectively. Higher cellulose content was estimated in U.lactuca (14.03 0.14%), followed by E.intestinalis (12.10 0.53%) and C.media (10.53 0.17%) least cellulose content was estimated in G. corticata (0.87 0.07%). Cellulose is a glucan present in green seaweeds, which can be easily hydrolysed by using enzyme and subsequently fermented to produce bioethanol. Green seaweeds are rich in cellulose content (>10%) [37,89,105] when compared to red and brown seaweed (2–10%).
3.1.3. Reducing sugar extraction using pretreatment methods
Chosen seaweeds composed of all the three types of algae (green, red and brown) with varied amounts of sugar and the complexity of the seaweed is reflected between structural and carbohydrate components [105,155,156]. It is therefore important to carefully choose the pretreatment process based on the biomass and an appropriate efficient pretreatment process to achieve a high yield of sugar for low energy input. The reducing sugar was determined using various pretreatment method as shown in Fig. 5. Pretreatment using boiling water yielded very low reducing sugar and it was observed that red and brown algae formed gel during this pretreatment process due to the presence of sugars like Agar, Carrageenan and Alginate which possess hydrocolloid properties [70,157,158]. It is seen that, liquid hot water pretreatment releases most of oligosaccharides [159]. U. lactuca biomass was subjected to four different pretreatments namely; ethanol organ solvent, alkaline, liquid hot water and ionic liquid treatments. Organo solvent and liquid hot water treatment produced highest glucan recovery of 80.8 g/100 g DW and 62.9 g/100 g DW respectively [160]. Sonication pretreatment also yielded lower reducing sugar, since sonication is most commonly used in extraction of lipid from microalgae [161], highest fatty acid composition of C16:0 and C18:1 was achieved in Spirogyra sp. employing sonication as cell disruptions technique [40]. Higher reducing sugar was extracted from all types of seaweeds during the dilute acid pretreatment. Dilute acid reduces the degree of polymerization resulting in recovery of 80–90% of hemicellulose exposing cellulosic fraction accessible to enzyme digestion [162]. Pre-treated Palmaria palmata, a red alga was exposed to acid hydrolysis, generated 218 mg/g of reducing sugar [133] and Grateloupia lithophila gave 191 mg/g of reducing sugar in the current study. Hence, it is feasible to obtain fermentable sugars with lower inhibitor concentration using extremely lower acid concentration [163]. Benefits of treating biomass with extremely low acid concentration simplifies downstream process such as neutralization and waste treatment. Reducing sugar of 14.7 g/L was obtained from E.intestinalis by dilute acid concentration of 75 mM H2SO4 [120]. Sequential acid hydrolysis was carried out to concentrate the sugar: hydrolysate of first cycle was utilized as hydrolyzing liquid for the 2nd cycle and up to 5th cycle [117] and generated 72 g/L of reducing sugar at the end of 5th cycle during 0.9 N H2SO4 hydrolysis from the seaweed granules. However, loss of liquid volume encountered in the hydrolysate that is attributed to the sorption loss by residue. Pulverized wet biomass of Gelidium amansii, red seaweed was subjected to continuous acid hydrolysis which yielded higher galactose and lower inhibitor concentration than batch reactor [126].
Utilization of red and brown seaweeds biomass for bioethanol production can lead to debate on hydrocolloid versus fuel affecting the existing multibillion hydrocolloid industry [89]. Therefore, for further processes of detoxification, enzyme hydrolysis and fermentation, two seaweeds Ulva lactuca and Enteromorpha intestinalis were selected as both the species satisfy the criteria of potential feedstock for bioethanol production such as; annual availability, carbohydrate rich biomass, producing higher reducing sugar concentration, ease of harvest by mechanical means, amenable to transplanting and reproducing prolifically in given environment [89].
3.1.3.1. Detoxification of acid hydrolysate.
Detoxification of inhibitors (HMF and LA) resulting from acid hydrolysis was carried out using activated charcoal [164], which removed 38.8% LA and 70.37% HMF. However activated charcoal also removed sugars 14.5% glucose and 20.3% galactose present in the hydrolysate [141]. In the current study, highest sugar removal of 63.5% and 52% was recorded from activated charcoal for U.lactuca and E.intestinalis acid hydrolysate respectively. Similarly, Ca(OH)2 was used for neutralizing the hydrolysate, which led to gypsum (calcium sulphate) formation, which was removed through filtration [117] also calcium ions catalyzes alkaline degradation of mono-saccharides in the over liming process, resulting in removal of fermentable sugars [141]. Similar results were observed in the current study, where Ca (OH)2 treatment removed sugar from U.lactuca (56.1%) and E.intestinalis (23.3%) acid hydrolysate. Least effect of sugar removal was recorded in Na2CO3 neutralization process (Table 6) U.lactuca (39.8%) and E.intestinalis (14.7%).
| |
Ulva lactuca |
Enteromorpha intestinalis |
| After treatment (mg/g) |
After treatment (mg/g) |
% Sugar removal |
After treatment (mg/g) |
After treatment (mg/g) |
% Sugar removal |
| Na2CO3 |
206.34 |
124.12 |
39.8 |
201.09 |
171.53 |
14.7 |
| NaOH |
|
113.78 |
44.9 |
|
153.88 |
23.3 |
| Activated Charcoal |
|
75.29 |
63.5 |
|
96.50 |
52.0 |
| Ca(OH)2 |
|
90.51 |
56.1 |
|
154.18 |
23.3 |
Table 6 Comparison of reducing sugar removal after neutralization process.
3.2. Enzyme hydrolysis
Enzyme hydrolysis of U.lactuca and E.intestinalis yielded lower reducing sugar, compared to pretreated biomass during the incubation period. In order to overcome recalcitrance in second generation feedstock, the biomass was pretreated prior to enzyme hydrolysis. The pretreatment removed the lignin and hemicellulose exposing the cellulose fibers for enzyme hydrolysis [64,66,99,165]. Pretreated biomass is then subjected to enzyme hydrolysis, where cellulase enzymes bind to cellulose and disintegrate it into simple sugars (glucose). Algal biomass in the current study was pretreated using dilute acid and then subjected to enzyme hydrolysis (Fig. 6a and b). Pre-treatment breaks down the crystalline cellulose structure, the major constituents in the biomass cell walls and maximize enzymatic conversion to reducing sugars [53,105, 155,166]. Despite longer incubation period for sugar release, enzyme hydrolysis is widely preferred due to the absence of any inhibitors formation [70].
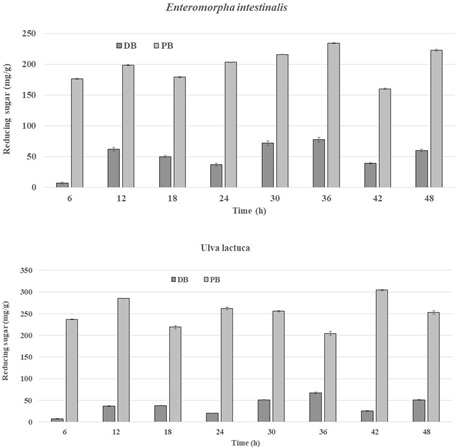 Fig. 6. a) Enzyme hydrolysis for direct biomass (DB) and acid pretreated biomass (PB) for E.intestinalis. b) Enzyme hydrolysis for direct biomass (DB) and acid pretreated biomass (PB) for U.lactuca.
Fig. 6. a) Enzyme hydrolysis for direct biomass (DB) and acid pretreated biomass (PB) for E.intestinalis. b) Enzyme hydrolysis for direct biomass (DB) and acid pretreated biomass (PB) for U.lactuca.
3.3. Bioethanol production from selected macroalgae
Production of bioethanol from carbohydrate rich biomass is economical and sustainable. However, investigations are focused on improving the yield of bioethanol from seaweeds using appropriate microorganisms that have capabilities to convert all sugars present in the seaweeds and achieve ethanol yield of 0.47 g/g reducing sugar [89, 167,168]. SHF and SSF process was carried out for E.intestinalis and U. lactuca feedstock.
3.3.1. SHF and SSF
Macroalgal feedstock E.intestinalis (EI) and U.lactuca (UL) were pretreated using 0.7 N and 0.5 N H2SO4 respectively. Seaweed hydrolysate was neutralized and then subjected to fermentation using CY and TY strain and conversion efficiency calculated assuming the sugar available in the hydrolysate are fermentable (Table 7). Higher ethanol yield of 1.63 g and 25.8% efficiency was recorded for EITY combination, whereas ULCY produced lower ethanol yield of 0.37 g achieving 12.1% efficiency. Co-fermentation of E.intestinalis hydrolysate yielded 0.8 g with 21.7% efficiency, whereas U.lactuca yielded 0.63 g/g achieving 20.4% efficiency [120]. Pretreated E.intestinalis to 75 mM H2SO4 and subjected to SHF and obtained ethanol of 8.6 g/L (0.86 g) achieving 30.5% efficiency at 48 h. At the end of fermentation 10 g/L of reducing sugar remained unutilized indicating presence of non-fermentable sugar not consumed by the yeast S.cerevisiae. U.fasciata enzyme hydrolysate was subjected to fermentation and 1.28 g of ethanol yield was produced achieving 88.27% efficiency indicating higher reducing sugar conversion efficiency and ethanol yield than those reported for red and brown seaweeds [169].
| Macroalgal feedstock |
Process |
Combinations |
Biomass (g) DW |
Initial sugar (g) |
Final sugar (g) |
Fermented sugar (g) |
Ethanol yield (g) |
Theoretical yield |
Efficiency (%) |
| Enteromorpha intestinalis |
SHF |
EICY |
50 |
22.5 |
10.71 |
11.79 |
1.40 |
6.01 |
23.4 |
| |
EITY |
|
|
10.13 |
12.37 |
1.63 |
6.31 |
25.8 |
| SCHF |
EITYCY |
|
|
15.09 |
7.41 |
0.82 |
3.78 |
21.7 |
| Ulva lactuca |
SHF |
ULCY |
|
19.2 |
13.17 |
6.03 |
0.37 |
3.07 |
12.1 |
| |
ULTY |
|
|
12.72 |
6.48 |
0.52 |
3.31 |
15.6 |
| SHCF |
ULTYCY |
|
|
13.10 |
6.10 |
0.63 |
3.11 |
20.4 |
CY: Cashew yeast; TY: Toddy yeast
Table 7 Fermentation of macroalgal feedstock by the process of separate hydrolysis and fermentation (SHF) and Separate hydrolysis and Co-fermentation (SHCF).
SSF of E.intestinalis produced 7.6 g/L with 26.9% efficiency, lower yield was attributed to the suboptimal temperature of 32 C which is unsuitable for enzyme hydrolysis [120]. In this study, SSF process (Table 8) produced higher efficiency when compared to SHF. The combination of EITY produced higher ethanol yield of 0.49g/g achieving 77.4% efficiency. Simultaneous saccharification and co-fermentation (SSCF) of E. intestinalis produced 0.27 g/g of ethanol with 41.8% efficiency whereas U.lactuca produced efficiency of 113% indicating fermentation of sugars other than glucan. Candida shehatea, Scheffersomyces stipitis (Pichia stipitis), and Pacchysolen tannophilus are the most promising yeast species for conversion of Xylose [112], which can be used in combination with S.cerevisiae in SSCF process to obtain higher ethanol yield [104]. subjected cellulosic residue of Kappaphycus alvarezii along with galactose synthetic medium to SSCF using single strain S.cerevisiae CBS1782 and recorded 64.3 g/L of ethanol indicating utilization of galactose sugar along with glucose. Co-fermentation of corn stover using S.cerevisiae and commercial strain TMB3400 yielded 40g/L or ethanol with 59% theoretical efficiency [74]. S.cerevisiae and S. stipitis were used for SSCF of Rice straw and produced 15.2g/L of ethanol [170].
| Macroalgal feedstock |
Process |
Combinations |
Biomass (g) DW |
Cellulose (g) |
Final sugar (g) |
Fermented sugar (g) |
Ethanol yield (g) |
Theoretical yield |
Efficiency (%) |
| Enteromorpha intestinalis |
SSF |
EICY |
20 |
1.8 |
0.34 |
1.46 |
0.25 |
0.74 |
33.3 |
| |
EITY |
|
|
0.56 |
1.24 |
0.49 |
0.63 |
77.4 |
| SSCF |
EITYCY |
|
|
0.52 |
1.28 |
0.27 |
0.65 |
41.8 |
| Ulva lactuca |
SSF |
ULCY |
|
1.7 |
0.54 |
1.16 |
0.33 |
0.59 |
54.9 |
| |
ULTY |
|
|
0.54 |
1.16 |
0.39 |
0.59 |
65.5 |
| SSCF |
ULTYCY |
|
|
0.66 |
1.04 |
0.60 |
0.53 |
113.0 |
CY: Cashew yeast; TY: Toddy yeast
Table 8 Fermentation of macroalgal feedstock by the process of Simultaneous Saccharification and Fermentation (SSF) and Simultaneous Saccharification and Co fermentation.
Seaweeds contain low amounts of polysaccharides composed of glucose. Production of ethanol, therefore, needs to be from carbohydrates including sulphated polysaccharides, sugar acids and sugar alcohols. However, inability of microorganisms in fermenting all sugars present in seaweeds into ethanol is a major drawback. Therefore, isolation of yeast strains to ferment both pentose (C5) and hexose (C6) sugars are vital for achieving higher ethanol yield.
However, recent studies are focusing on non-controversial cellulosic residue following extraction of hydrocolloid from seaweed biomass [60]. But the lower cellulose content of residue prevents it from being a viable feedstock option considering the emergent demand for bioethanol. This necessitates the selection of seaweed species with higher cellulose content together with higher growth rate for sustainable bioethanol production [89,171]. Red algae has more agar and carrageenan and brown algae has more algin and lower cellulose compared to green seaweeds, that have cellulose making up to 70% of their dry weight (as Cladophorales and Ulvales) [176].
It is evident from the experimental results, that green algae are apt as potential feedstock due to higher carbohydrate and cellulose contents. Higher growth rates have been recorded for green seaweeds ranging from 19.15 to 24.25% when compared to red macroalgae (3–8%) [89, 177,178]. Green seaweeds have production potential that is 2–20 times that of conventional terrestrial energy crops [179]. Red and brown seaweed’s lower cellulose content of residue and lower growth rate prevents it from being a viable feedstock to meet the growing demand for bioethanol. Comparison of ethanol yield from other potential macroalgal feedstock are represented in Table 9.
| Seaweed species |
|
Reducing sugar g/l |
Ethanol yield g/g |
%Theoretical yield |
References |
| Green Seaweeds |
Chaetomorpha media |
27.79 |
0.057 |
10.15 |
* |
| Ulva fasciata |
21.82 |
0.43 |
83.66 |
[89] |
| Ulva lactuca |
1.12 |
0.39 |
77.03 |
[94] |
| Ulva lactuca |
35.43 |
0.23 |
45.62 |
* |
| Ulva pertusa |
43 |
0.43 |
84.36 |
[70] |
| Ulva pertusa |
26 |
0.48 |
93.51 |
[97] |
| Enteromorpha intestinalis |
48.96 |
0.25 |
49.37 |
* |
| Enteromorpha intestinalis |
45.56 |
0.21 |
41.74 |
[120] |
| Red Seaweeds |
Gelidium elegans |
49 |
0.38 |
73.63 |
[70] |
| Gelidium amansii |
43.5 |
0.47 |
92.40 |
[108] |
| |
42.2 |
0.38 |
74.51 |
[126] |
| |
7.93 |
0.42 |
82.34 |
[172] |
| |
7.93 |
0.48 |
93.46 |
|
| |
7.93 |
0.48 |
93.46 |
|
| Gracilaria verrucosa |
34.63 |
0.43 |
84.31 |
[60] |
| |
19.70 |
0.43 |
84.31 |
[110] |
| Gracilaria corticata |
57.90 |
0.01 |
0.98 |
* |
| Gracilaria salicornia |
13.8 |
0.079 |
15.49 |
[173] |
| Gracilaria sp. |
11.46 |
0.42 |
82.35 |
[132] |
| Grateloupia lithophila |
44.45 |
0.09 |
17.85 |
* |
| Gelidium pusillum |
36.96 |
0.04 |
8.49 |
* |
| Eucheuma cottonii (Kappaphycus alvarezii) |
11 |
0.45 |
89.13 |
[88] |
| |
81 |
0.45 |
88.24 |
[104] |
| |
20.4 |
0.21 |
41.18 |
[164] |
| |
51.9 |
0.42 |
82.36 |
[117] |
| Palmaria palmata |
21.84 |
0.173 |
33.92 |
[133] |
| Brown Seaweeds |
Alaria crassifolia |
67.20 |
0.38 |
74.40 |
[70] |
| Laminaria hyperborea |
30 |
0.43 |
84.31 |
[114] |
| Padina tetrastromatica |
17.9 |
0.01 |
2.53 |
* |
| Sargassum sagamianum |
25.9 |
0.386 |
75.69 |
[116] |
| Sargassum sagamianum |
2.55 |
0.89 |
174.55 |
[174] |
| Sargassum sagamianum |
19.8 |
0.35 |
69.32 |
[125] |
| Sargassum ilcifolium |
27.04 |
0.05 |
9.60 |
* |
| Sargassum fluvellum |
9.6 |
|
|
[54] |
| Saccharina japonica |
6.72 |
0.343 |
67.25 |
[163] |
| Saccharina japonica |
34 |
0.41 |
80.74 |
[103] |
| Saccharina japonica |
45.6 |
0.17 |
33.11 |
[52] |
| Saccharina japonica |
75 |
0.34 |
67.45 |
[116] |
| Saccharina latissima |
35 |
0.45 |
88.24 |
[36] |
| Undaria pinnatifida |
42.9 |
0.22 |
43.05 |
[120] |
| Undaria pinnatifida |
20 |
0.144 |
28.24 |
[107] |
| Seagrass |
Zosteria marina |
92 |
0.059 |
11.5 |
[175] |
*Current work.
Table 9 Comparison of reducing sugar, ethanol yield and percent theoretical yield from various macroalgal biomass.
3.3.2. Multivariate analysis of process condition for bioethanol production
Downstream process of bioethanol production from macroalgal biomass is dependent on various factors such as dilute acid pretreatment conditions (acid concentration, temperature, incubation time), enzyme hydrolysis conditions (pH, temperature and incubation time), reducing sugar and fermentation conditions (temperature and incubation time). In order to understand influence of each of these factors in ethanol production, multivariate regression analysis (Table 10) was performed and the probable relationship is given by equation (2). Overall this model explains 97.3% variation in the data
Y=-2.75-4.09(X1)+0.12(X2)-0.13(X3)+10.61(X4)-0.31(X5)-0.04(X6)+0.32(X7)-1.23(X8)-0.12(X9)------------>(2)
Where, Y: Ethanol g/L, X
1: Acid concentration (mM), X
2: Acid hydrolysis Temperature (°C), X
3: Acid hydrolysis incubation time (min), X
4: buffer pH, X
5: Enzyme hydrolysis temperature (°C), X
6: Enzyme hydrolysis incubation time (h), X
7: Reducing sugar in (g/L), X
8: Fermentation temperature (°C), X
9: Fermentation time (h)
This analysis highlights that process conditions such as; acid pretreatment temperature, buffer pH and reducing sugar concentration affects the ethanol yield (Eq. (2)). The increase in hydrolysis temperature during acid catalyzed reaction would also enhance the sugar production and decomposition [180,181]. Pretreatment temperature plays a vital role in sugar release as higher temperatures often leads to the formation of inhibitor, which is detrimental to the fermentative microorganisms leading to reduced ethanol yields [69,181]. Enzymes consists of ionic groups on their active sites which varies along with the pH resulting in changes in the activity of the enzyme, its structure, reaction rate and the product formation [182]. Therefore, pH of the reaction medium or buffer needs to be optimum for higher enzyme activity [183, 184]. Fermentation process is slower in low sugar medium, whereas the process increases in the medium containing 15–20 g/L of sugar and remains stable up to 200 g/L. Medium with 200 g/L to 300 g/L of sugar concentration lowers the growth of yeast microorganisms [185–187]. However, higher concentration of fermentable sugars yield higher ethanol, whereas non-fermentable sugars (pentose) can affect the fermentation yield due to lack of transport system in yeasts.
| Process |
Process condition |
Coefficients |
Standard Error |
| Dilute acid pretreatment |
Concentration |
-4.09 |
1.14 |
| Temperature |
0.12 |
0.29 |
| Incubation Time |
-0.13 |
0.67 |
| Enzyme hydrolysis |
pH |
10.61* |
2.44* |
| Temperature |
-0.31 |
0.16 |
| Incubation Time |
- 0.04 |
0.02 |
| Reducing sugar |
0.32* |
0.04* |
| Fermentation process |
Temperature |
- 1.23 |
2.09 |
| Incubation Time |
0.12* |
0.02* |
*p <0.05.
Table 10 Coefficients and SE values of multivariate regression analysis.

