|
DISCUSSION
The results of this study show that in the Western
Ghats, some ant species and even some functional
groups establish their nests at locations that can
be clearly distinguished using LISS-derived NDVI.
However, the discriminative success of NDVI was
limited to ant species that nest in areas with low
NDVI, i.e. in relatively sparse vegetation. The
reason for the failure of NDVI to distinguish ant
nesting locations at the species (and functional
group) level when they are located in dense vegetation is probably that in these cases, ant
species do share similar habitat in terms of
greenness and / or biomass: H. saltator and L.
processionalis are specialist predators on other
arthropods and known to inhabit deciduous and
evergreen forests; O. smaragdina and C.
taprobanae are arboreal ants that are dominant in
humid tropical regions, and P. diversus is a cryptic
species that nests and forages within soil and leaf
litter (Figs. 3, 4). To distinguish nesting sites of
such species it might therefore be necessary to
use additional variables.
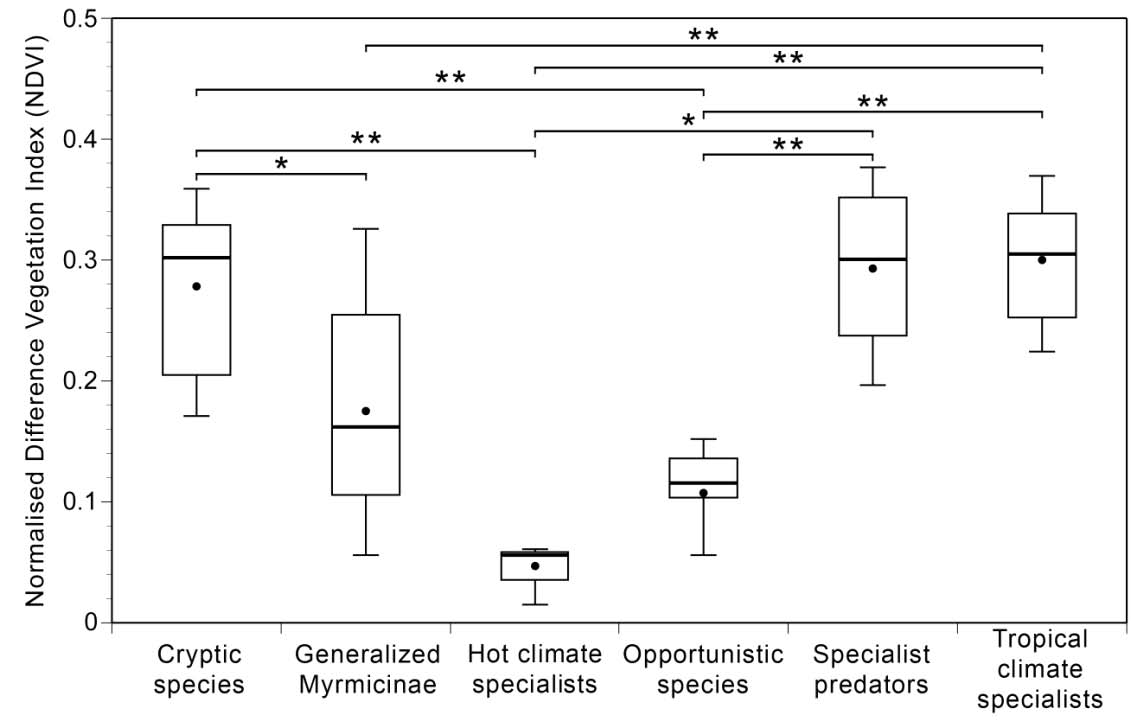
Fig. 3 : Box plot illustrating the variation of NDVI at nest locations of six functional groups. Each box shows
upper and lower quartiles along with 90th and 10th percentiles (whiskers), median (thick line) and mean (filled
circle). NDVI values are shown on y-axis. The six functional groups are shown on x-axis. Note: Pachycondyla
rufipes has not been included in this functional group analysis. NDVI at the nest sites of the Cryptic Species
(n=14), Generalised Myrmicinae (n=59), Hot Climate Specialists (n=6), Opportunistic Species (n=22), Specialist
Predators (n=14) and Tropical Climate Specialists (n=21) were significantly different (P < 0.001). Pairs that
are significantly different are highlighted with * or ** (P < 0.01 and P < 0.001 respectively, Tukey test).
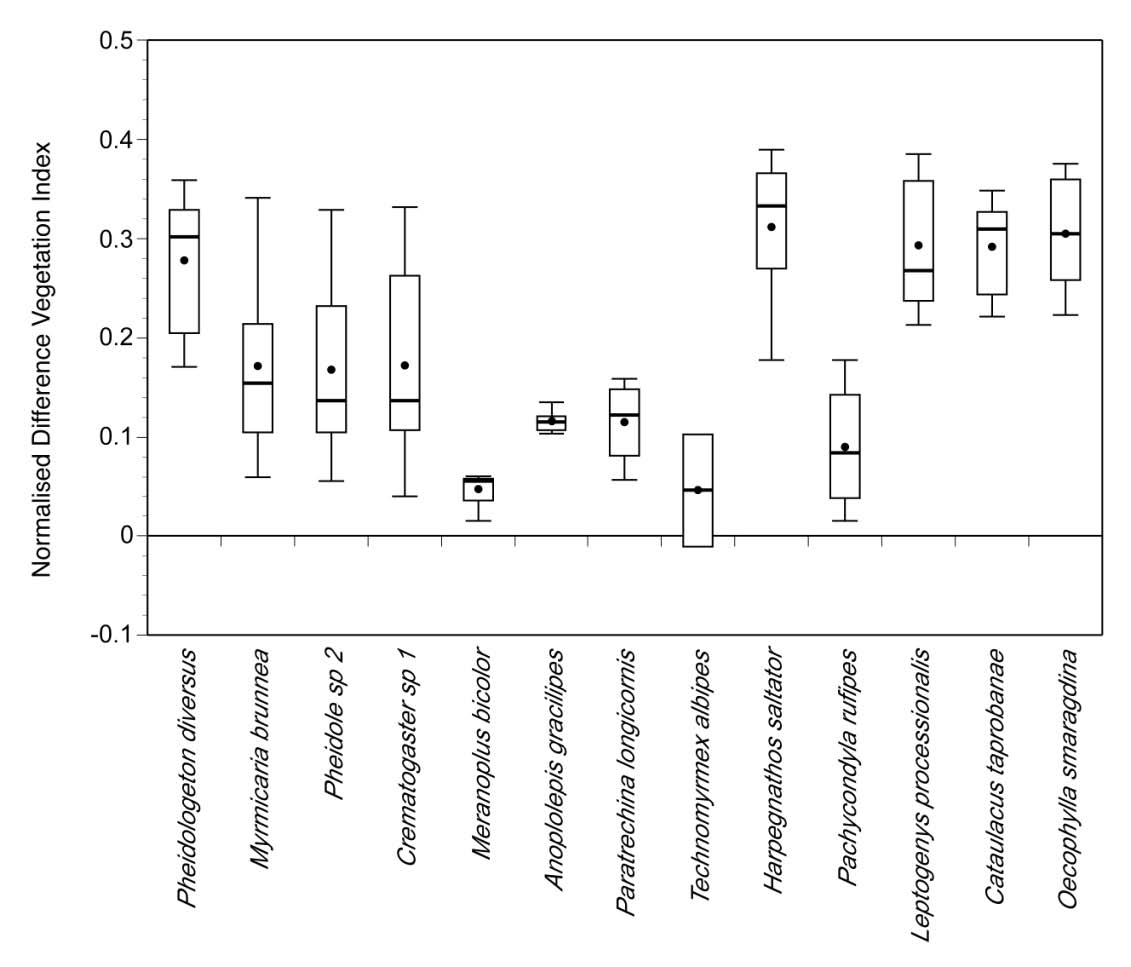
Fig. 4 : Box plot illustrating the variation of NDVI at nest locations of 13 species. Each box shows upper and
lower quartiles along with 90th and 10th percentiles (whiskers), median (thick line) and mean (filled circle). NDVI
values are shown on y-axis. The different species are shown on x-axis. NDVI at the nest sites of Anoplolepis
gracilipes (n=9), Paratrechina longicornis (n=8), Technomyrmex albipes (n=5), Cataulacus taprobanae (n=8),
Oecophylla smaragdina (n=13), Harpegnathos saltator (n=7), Pachycondyla rufipes (n=11), Leptogenys
processionalis (n=7), Crematogaster sp. 1 (n=21), Pheidole sp. 2 (n=25), Myrmicaria brunnea (n=13),
Pheidologeton diversus (n=14) and Meranoplus bicolor (n=6) were significantly different (P < 0.001). Tukey
test revealed significant differences (P < 0.01) between the following pairs: M. brunnea vs O. smaragdina;
Crematogaster sp. 1 vs Pheidole sp. 2; Crematogaster sp. 1 vs O. smaragdina; Crematogaster sp. 1 vs P.
diversus; Pheidole sp. 2 vs H. saltator; Pheidole sp. 2 vs L. processionalis; Pheidole sp. 2 vs C. taprobanae;
Pheidole sp. 2 vs O. smaragdina; Pheidole sp. 2 vs P. diversus; A. gracilipes vs H. saltator; A. gracilipes vs L.
processionalis; A. gracilipes vs C. taprobanae; A. gracilipes vs O. smaragdina; T. albipes vs H. saltator; T.
albipes vs L. processionalis; T. albipes vs C. taprobanae; T. albipes vs O. smaragdina; T. albipes vs P. diversus;
P. longicornis vs L. processionalis; P. longicornis vs C. taprobanae; P. longicornis vs O. smaragdina; P. longicornis
vs P. diversus; P. longicornis vs H. saltator; H. saltator vs P. rufipes; H. saltator vs P. diversus; H. saltator vs M.
bicolor; P. rufipes vs L. processionalis; P. rufipes vs C. taprobanae; P. rufipes vs O. smaragdina; P. rufipes vs P.
diversus; L. processionalis vs M. bicolor; C. taprobanae vs M. bicolor; O. smaragdina vs M. bicolor and P.
diversus vs M. bicolor.

Fig. 5 : Profile view of Pachycondyla rufipes (Jerdon), an ant that establishes terrestrial nests under canopy gaps
in moist deciduous and evergreen forests. Collected at Sharavathi River Basin, Shimoga, Western Ghats, India.
A potential limitation of this study is that
our interpretation of the relationship between ant
nest sites and habitat type was based on nest-site
NDVI only. Although we complemented this with
a correlation analysis of NDVI and the pre-defined
habitat types, a direct assessment of speciesspecific
nest site frequency vs. habitat type is
missing. Regardless of this shortcoming, our
purely NDVI-based analysis results match the
documented habitat preferences of species
inhabiting areas with less dense vegetation quite
well: invasive species such as A. gracilipes and P.
longicornis along with T. albipes were abundant
in scrub jungles and acacia plantations that have
low NDVI (Fig. 4). All three species are
Opportunists that exhibit unspecialised food and
niche requirements, are poorly competitive, and
are dominant in disturbed habitats (Andersen
1995). The seed harvesting ant M. bicolor was
abundant in regions with low NDVI such as scrub
jungles. On the other hand, a particular wide range
of NDVI niches were occupied by the ubiquitous
species of the myrmicine community
(Crematogaster sp. 1, Pheidole sp. 2 and M.
brunnea) that do not have highly specific niche
requirements (Fig. 4).
A surprising finding was that P. rufipes, a
specialist predator on termites (Narendra & Kumar 2006), built nests at sites with low NDVI despite
the fact that this species was collected only from
deciduous and evergreen forests (Fig. 4). Similar
observations on the niche occupied by this ant
species have been reported (Narendra & Kumar
2006; Narendra et al. in review). NDVI at the
collection sites of P. rufipes was similar to that in
the scrub jungles (Fig. 3), a habitat from which
this species was never collected. And although
our validation fieldwork confirmed the initial
observation that both nesting sites and foragers
of P. rufipes coincided with canopy gaps of
deciduous and evergreen forests, the question
remains: why is P. rufipes found in canopy gaps?
Individually foraging ants rely on direct-ional
information gathered either from celestial cues
(Wehner 2001) or from landmarks present in the
foraging environment (Fukushi 2001; Narendra
2007). Solitary foraging ants are well known for
their ability to return to the nest by matching the
previously seen views (Wehner & Räber 1979;
Narendra et al. 2007). In fact a congeneric species
of P. rufipes, Pachycondyla tarsata (Fabr.) (previously
known as Paltothyreus tarsatus) uses the
contrast available in the canopies (Hölldobler 1980)
to match its previously acquired image to its
current images to return to the nest. In the landmarkrich
habitats of P. rufipes it is quite unlikely for
individual trees to act as a beacon and utilisation
of celestial cues may be hindered. It is perhaps
because of this that P. rufipes colonises canopy
gaps, a micro-niche that would enable the ants to
forage using information derived from both
canopies and sky.
The example of P. rufipes demonstrates the
potential of high-resolution remotely sensed NDVI
data in delineating preferred nesting sites for
species whose habitat preferences are clearly
different from those of other ant species in the
study area, in terms of both density and geometry
of the local vegetation. It does not however shed
much light on the potential of NDVI as a predictor
variable to model ant species distributions, as this
would require validating results for each species
at random locations from anywhere in the study
area and not only from within the high-probability
range. Also, if NDVI was to be tested as predictor
variable for presence of P. rufipes nests, we would
recommend to not only employ the absolute value
but also a derived variable that quantifies the
difference in NDVI between neighbouring pixels,
i.e. taking into account the “canopy gap” as an
argument for habitat suitability. We emphasise that
our study did not intend to validate the pre-defined
habitat categories using remotely sensed NDVI;
instead, we merely assessed the correlative
strength between these two at the selected nest
sites. It would be interesting however, to use the
LISS-derived NDVI image to establish habitat
types for the whole study area (e.g. by means of a
supervised classification) and then explore the
relationship between ant nest site locations and
habitat type at both the species and the functional
group level.
We conclude that LISS-derived NDVI has
considerable value in deriving the nest site
locations of some ant functional groups and even
at species level, especially regarding ant species
that belong to the functional groups Hot Climate
Specialists and Opportunists. These results are
encouraging for decision-makers dealing with
invasive species, which are often opportunistic.
Officers in the land-use and conservation sector
who need to monitor the effects of intensifying
human land-use and climate change stand to
benefit as well, since many ant species are
considered reliable indicators for ecosystem
change
Top |

