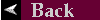LIMNOLOGY
Essentials
BIOLOGICAL
FACTORS
PLANKTON COMMUNITY:
Organisms that passively drift, maintained in suspension by water current, or float or swim weakly comprise the plankton. They include heterotrophic bacterioplankton, photosynthetic phytoplankton and zooplankton.
Planktons refer to microscopic aquatic plants or animals having little or no resistance to water current and living free floating and suspended in open or “pelagic waters”. Planktonic plants are called as phytoplanktons and animals as zooplanktons. The phytoplankton (microscopic algae) occur as unicellular, colonial or filamentous forms. Many are photosynthetic and are grazed by zooplankton. The algae of the open water of lakes and streams consist of diverse assemblage of all major taxonomic groups. Many of these forms have different physiological requirements and differ in their response to light, temperature and concentration of nutrients.
Populations of algae and the animals that feed on them are lower in oligotrophic lakes because of low nutrient concentrations. Thus the water remains clear. Decay of relatively small amounts of organic matter in oligotrophic lakes does not completely deplete the hypolimnetic supply of dissolved oxygen. Therefore the lack of oxygen does not restrict animals from living in the hypolimnion of oligotrophic lakes.
These lakes have two types of fishery with warm and cool water in the epilimnion and metalimnion and fish present in the cold oxygen rich portion of the hypolimnion. Coldwater fishery is very sensitive to increased inputs of organic matter. Algae or macrophytes grow thickly in eutrophic lakes where light penetrates only a short distance and nutrients below the depth are not assimilated. Phosphorus is the limiting nutrient in freshwater lakes, as the plants deplete all the available phosphorus before they deplete other nutrients. In a hyper eutrophic lake, algae may become abundant. When abundant phosphorus is available in a lake, nitrogen may become limiting. In such lakes, certain species of blue green algae that can fix atmospheric nitrogen have a clear competitive advantage and frequently become dominant. Eutrophic lakes show wide seasonal changes in their biological and chemical conditions. Because of organic matter produced in these lakes, the decay rate is high in the hypolimnion, causing oxygen to deplete. Therefore, eutrophic lakes frequently show a complete loss of dissolved oxygen below the thermocline during summer.
Phytoplankton:
Phytoplanktons (Figs. 4 to 8) are of many species, mostly oxygen-evolving prokaryotic blue green algae (cyanophyta) and prochlorophyta and eukaryotic algae. The eukaryotic groups comprise Cryptophyceae (cryptophytes), Dinophyceae (dinoflagellates), Chlorophycota (green algae), Euglenophycota (euglenoids), Bacillariophyceae (diatoms), Chrysophyceae and Haptophyceae (yellow-green or golden-yellow algae). Most of the phytoplanktons are denser than water. Some have flagella, whose movement may counter the tendency to sink. Non-flagellated types have evolved cell or colony shapes that decrease the rate of sinking. Phytoplanktons satisfy their carbon and energy needs through photosynthesis. Some need at least one preformed organic compound, usually a vitamin, such as cyanocobalamin (B12), thiamine or biotin. In general, blue green algae, diatoms and desmids do not require additional organic compounds, but many algae of other groups are able to take up simple organic compounds heterotrophically.
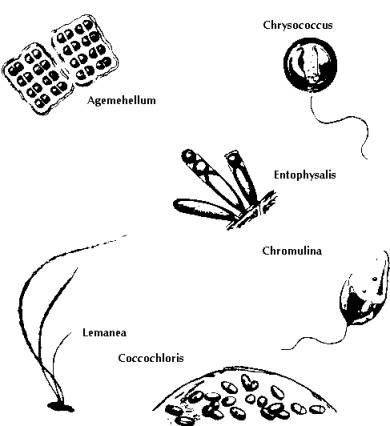
Fig 4: Clean Water Algae
Phytoplanktons need about 20 elements for growth, but only carbon, nitrogen and phosphorous are likely to limit the growth rates. All these elements are present in water at lower concentrations than that required by the cell. Hence, energy-requiring mechanisms involving enzymes are needed to concentrate them in cells. In 1934, Redfield suggested that cells which had a balanced nutrient would have carbon:nitrogen:phosphorous ratio of 106:16:1, and severe departures from this indicate nutrient limitation. In fresh waters due to their small size, local variations in loading rates especially those caused by human activity disrupts the ecological balance. The imbalance of artificially altered phosphorous and nitrogen loading may create scarcity, for e.g., heavy phosphorous loading may induce nitrogen scarcity. Freshwater phytoplankton communities change according to human influence. The appearance of phytoplanktons depends on the availability of nutrients, degree of stratification and incidence of grazing. The phytoplankton of an oligotrophic lake is likely to be of small organisms with high surface to volume ratios. Eutrophic waters may be able to sustain greater proportions of larger organisms. The blue green algal blooms are a common phenomenon during summer and give rise to neurotoxins and hepatotoxins. Neurotoxins are often anatoxins (secondary amine alkaloids) or saxitoxins (cyclic polypeptides). Hepatotoxins come in over 45 variants, each a related cyclic polypeptide. There is increasing evidence that these toxins may be passed into food webs and hence may have widespread effects.
A primary algal characteristic is the presence of photosynthetic pigments – the chlorophylls, carotenoids and biliproteins. Chlorophyll – a is the primary photosynthetic pigment of all photosynthetic organisms other than photosynthetic sulphur bacteria. Chlorophyll – b, although common in higher plants is found only in the green algae, euglenophytes and certain minor groups. Chlorophyll – b is the light gathering pigment that transfers absorbed light energy to chlorophyll - a. Chlorophyll – c consists of three spectrally distinct components and is an accessory pigment. Chlorophyll - d is a minor pigment found in certain red algae. Chlorophyll – e is present in certain xanthophycean algae. Among the many carotenoids of algae, the carotenes are linear unsaturated hydrocarbons, and the xanthophylls are oxygenated derivatives of carotenes. The light energy absorbed by carotenoids and biliproteins is transferred to chlorophyll – a, leading to fluorescence and excitation of chlorophyll - a molecules. b - carotene is the most widely distributed of the carotenes and is replaced by a - carotene only in certain green algae and in the cryptophyceae. The biliproteins are water-soluble, pigment-protein complexes in the cyanobacteria.
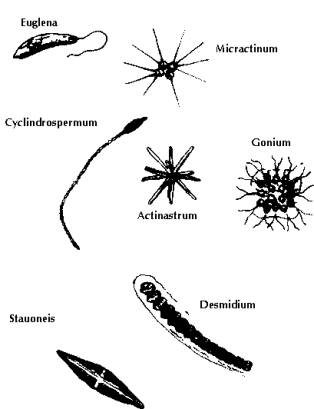
Fig 5: Polluted Water Algae
Cyanobacteria: The cyanobacteria (blue-green algae), classified in the division Cyanophyta or Myxophyceae, are true bacteria with a simple prokaryotic cell structure. Prokaryotic cells lack certain membranous structures, including a nuclear membrane, mitochondria and chloroplasts. The cells do contain cellular inclusions, which in some cases assume similar functions to eukaryotic organelles such as pigment-bearing lamellae, a plasma membrane and a nucleus containing chromosomal material. Cyanobacteria are differentiated from other bacteria by the presence of chlorophyll - a, which is common to chloroplasts of eukaryotic algae and higher plants. Cyanobacteria are also capable of using water as electron donor in photosynthesis, which is more advanced than bacterial photosynthesis. Thus, cyanobacteria possess both the enzymatic and pigment capability to photosynthesise under oxygenated conditions as in higher plants. Cyanobacteria occur as unicellular, filamentous organisms in mucilaginous sheaths as colonies or as individuals. Most of the planktonic cyanobacteria consist of the coccoid family Chroococcaceae (e.g., Microcystis, Coelosphareium and Coccochloris) and filamentous organisms (Planktothrix, Limnothrix and Tychonema), Nostocaceae (Anabena, Aphanizomenon and Nodularia) and Rivulariaceae (e.g., Gleotrichia).
Cyanobacteria contain differentiated cells called as heterocysts. Heterocysts can occur in many filamentous forms but are absent from those in oscillatoria; they are major sites of nitrogen fixation. Heterocysts lack phycobilins, which are the primary light absorbers in cyanobacteria and lack oxygen evolving photosystem and show higher reducing activity. The waxy layers in the envelope of the heterocyst restrict the rate of oxygen diffusion from outside, but sufficient nitrogen gets through to support nitrogen fixation. The internal environment of the heterocyst is conducive for nitrogen fixation, since the nitrogenase enzyme (present inside the heterocyst) is inactivated by the presence of oxygen. Organic carbon from the adjacent cells is transferred into the heterocyst and used as an energy source in the nitrogen fixation.
Fig 6: Algae found attached to a Substrate
Green algae: The chlorophyta are an extremely large and morphologically diverse group of algae. Most of the planktonic green algae belong to the orders Volvocales (e.g., Chlamydomonas, Sphaerocystis, Eudorina, and Volvox) and Chlorococcales (e.g., Scenedesmus, Ankistrodesmus, Selanastrum and Pediastrum). Asexual reproduction by vegetative division is common to most of the green algae but is lacking in most of the Chlorococcales and Siphonales. Cell division in colonial species results in enlargement of the colony; new colonies are formed by the fragmentation of the colony. Sexual reproduction in the green algae is diverse. In the simplest case of isogamy and anisogamy, the flagellated male and female gametes are morphologically similar in size and structure or the female is slightly larger than the male. Among specialised algae, oogamy is common where the union occurs between a large nonflagellated female gamete and a small flagellated male antherozoid. Some green algae are sexually monoecious (gametes are derived from the small cell) and others are dioecious (male and female cells are derived from different cells). Whereas planktonic Volvocales and Chlorococcales are ubiquitous in distribution among waters of differing salinity within the limnological range, the distribution of most species of desmids is limited to low concentration of the divalent calcium and magnesium.
Yellow-Green algae: The xanthophyceae are unicellular, colonial or filamentous algae that are characterised by conspicuous amounts of carotenoids in comparison to chlorophylls that have predominately yellow-green colouration. All the motile cells have two flagella, one of which is smooth and longer than the other “hairy” flagellum. A cell wall is absent but when present contains large amounts of pectin. Asexual reproduction is usually through fission by the formation of zoospores. Most xanthophycean algae are associated with substrata and many are epiphytic on larger aquatic plants. A few members are planktonic and include common genera such as Chlorobotrys, Gleobotrys and Gloeochloris.
Fig 7: Plankton and other Surface Water Algae
Golden Brown algae: The chromatophores of the Chrysophyceae often yield a golden brown colouration because of the dominance of b-carotene and specific xanthophyll carotenoids in addition to chlorophyll-a. Most of the chrysophycean algae are unicellular; a few are colonial and rarely filamentous. Many species lack a cell wall and are covered by cytoplasmic membrane whereas some possess a cell surface covered by siliceous or calcareous plates or scales. Vegetative reproduction by longitudinal cell division is most common. The unicellular species with a single flagellum include Chromulina, Chrysococcus and Mallomonas and the larger colonial forms comprise of Synura, Chrysophaerella, Uroglena and Dinobryon. Several species of Chrysophyceae algae (particularly Dinobryon) supplement photosynthesis with phagotrophy. These algae obtain an appreciable amount of their energy and nutrients by ingesting bacteria. Although the rates of water clearance are small (e.g., 0.00014 ml/day) large populations of Dinobryon can collectively clear water of bacteria at rates that compete effectively with those of ciliates, rotifers and crustaceans.
Diatoms: The Bacillariophyceae are characterised by silicified cell walls and can be both colonial and unicellular. The group is mainly divided into centric diatoms (Centrales) having radial symmetry and the pinnate diatoms (Pennales), which exhibit bilateral symmetry. The cell wall or frustule of diatoms consists of two lid like valves, one of which fits within the other. The valves of the pinnate diatoms exhibit various areas of cell thickenings and dilations. In some species a slit, termed raphe, traverses all or part of the cell wall, in others, a depression in the axial areas of the cell wall termed as pseudoraphe is found. The four major groups of pinnate diatoms are differentiated on the basis of these structures: a) Araphidineae (possessing a pseudoraphe, e.g., Asterionella, Diatoma, Fragileria and Synedra); b) Raphidioidineae in which a rudimentary raphe occurs at the cell ends (Actinelia and Eunotia); c) Monoraphidineae which have a raphe on one valve and a pseudoraphe on the other. (e.g., Achnanthes and Cocconeis) and d) Biraphidineae, in which the raphe occurs on both valves (Amphora, Cymbella, Gomphonema, Navicula, Nitzschia, Pinnularia and Surirella). Vegetative reproduction by cell division is the most common mode of multiplication. Sexual reproduction occurs only when the cells reach a minimum critical size and correct environmental conditions (often species-specific), which may include combinations of light, temperature, nutrients, trace metals, organic growth factors etc.
Fig 8: Taste and Odour Algae
Cryptomonads: Most of the cryptophycean algae are unicellular and motile. The planktonic members belong to the Cryptomonadineae (e.g., Cryptomonas, Rhodomonas, and Chroomonas). The one or usually two chromatophores contain a variety of pigments. Reproduction is by longitudinal division. Cryptomonad algae and other cryptomonad like microflagellates occur in almost all lakes, regardless of the trophic state. Characteristics of cryptomonads include intermittent numerical dominance, high nutritional quality, short turnover times and ability to grow and reproduce at low intensities of light.
Dinoflagellates: The dinoflagellates are unicellular flagellated algae, many of which are motile. The few species do not have a cell wall (e.g., Gymnodinium of the Gymnodiniales), most develop a cell wall that bears large spines and elaborate cell wall processes (e.g. the Peridiniales - Ceratium, Glenodinium, Peridinium). The cell surface has transverse and longitudinal furrows that connect and contain the flagella. The movement of flagella create water currents providing for weak locomotion and the disrupting of chemical gradients at the cell surface. Although sexual reproduction occurs, asexual reproduction by the formation of aplanospores dominates.
Euglenoids: Although the euglenoid algae (Euglenophyceae) are relatively large and diverse, few species are truly planktonic. Almost all euglenoids are unicellular, lack a distinct cell wall and possess one, two or three flagella that arise from an invagination in the cell membrane. Reproduction occurs by longitudinal division of the motile cell. Nutrition is by photosynthesis but some are phagotrophic. Nutrition is supplemented by the uptake of dissolved organic compounds. Ammonia and dissolved organic nitrogen compounds are the dominant sources of nitrogen among most euglenoid algae. Their development occurs most often in seasons, strata or lake systems in which concentrations of ammonia and especially dissolved organic matter is high.
Brown and Red algae: The brown algae (Phaeophyta) are mostly filamentous or thalloid algae, which are essentially marine but a few genera are found in fresh water and are attached to substrata, such as rocks. The red algae (Rhodophyta) are also very sparsely spread in fresh water and are not planktonic. The thalloid or filamentous species (e.g., Batrachospermum) are all restricted to fast flowing streams of well-oxygenated water.
Some important factors affecting the growth and succession of phytoplankton are a) light and temperature; b) buoyancy regulation - means of remaining within the photic zone by alterations of sinking rates; c) inorganic nutrient factors; d) organic micronutrient factors and interactions of organic compounds with inorganic nutrient availability and e) biological factors of competition for available resources and predation by other organisms.
Interactions
of light and temperature frequently result in one of several possible vertical
profiles of photosynthesis in planktonic communities. In some cases there is a
zone of maximum photosynthetic rates at light saturation, which overlies a zone
of near exponential decline of rates with increasing depth. The depth at which
maximum photosynthesis occurs varies with transparency of the water, governed by
the concentration of dissolved and particulate organic matter and abiotic
turbidity. There is great diversity in the tolerance to variations in
temperature among the algae. The minimum temperatures at which photosynthesis
can occur depend upon the algal species. For many diatoms, the critical
temperature is about 5°C,
for others about 15°C
and in many green algae and cyanobacteria, even higher temperatures are needed.
Zooplankton:
Planktonic animals are dominated by four major groups a) protists that include protozoa and heterotrophic flagellates, b) rotifers and two subclasses of the crustacea, c) cladocera and d) copepoda.
Protozoa and other protists (Fig. 9): The free–living protists include the ciliates, most of the flagellates and sarcodines. These protists are eukaryotes (usually unicellular) and possess a nuclear envelope, eukaryotic ribosomal RNA and endoplasmic membranes. Maximum growth rates are found when food availability is not a constraint; under these conditions populations generally increase directly with temperature and decrease as body size increases. Many protists develop cysts under harsh environmental conditions. They are important microbial consumers and have major functions in organic carbon utilisation and nutrient recycling.
a. Flagellates: These are the most abundant components of the protozooplankton. Common forms include dinoflagellates (Ceratium and Peridinium), chrysomonads (Dinobryon, Mallomonas and Synura), euglenids (Euglena), volvocids (Volvox and Eudorina), choanoflagellates (Astrosiga) and the diverse and large group of heterotrophic flagellates. The two main groups on the basis of size are (a) heterotrophic nanoflagellates (below 15 mm) and b) large heterotrophic flagellates (within a range of >15-200 mm). Among both temperate and subtropical lakes, dinoflagellates tend to increase in dominance as the pH of the water decreases and can dominate in acidic lakes.
b. Ciliates: Of the three major groups of ciliate genera, the oligotrichs (Strombidium and Halteria) are found in tropical lakes. The tintinnid ciliates (Order Choreotrichida: Tintinnidium, Tintinnopsis and Codonella) are widely distributed in temperate to tropical regions. Haptorid ciliates (Askenasia and Mesodinium) are similarly broadly distributed and abundant. Ciliates supplement nutrition by photosynthesis but most are holozoic and feed on bacteria, algae, particulate detritus and other protists. Ciliates tend to be more significant components of the zooplankton of eutrophic lakes.
c. Sarcodines: Sarcodine protozoa are poorly found in the zooplankton of fresh waters. Even in eutrophic lakes, sarcodines occur in modest abundance.
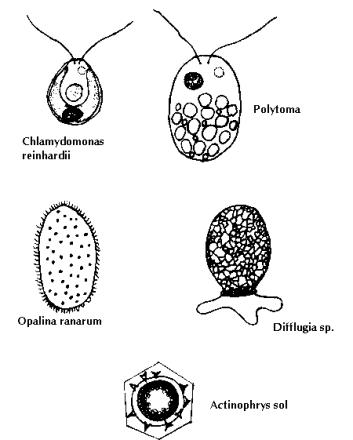
Fig 9: Protozoans
Populations of protists often develop in water strata greatly reduced in or devoid of oxygen in which bacterial populations tend to be dense, such as in lower portions of the hypolimnia of the meromictic lakes. Ciliates and flagellates are typically distributed in the upper water column in relation to the depth distribution of light. The euphotic zone is shallower among eutrophic lakes (often <5m) but extends much deeper in less productive lakes. Some flagellates such as Ceratium can position themselves in areas of maximum light availability, nutrient and oxic conditions. Seasonal variations are generally noticed in flagellate and ciliate abundance. Cryptomonad and other microflagellates are numerically dominant across a spectrum of oligotrophic through hypereutrophic lakes. Generally ciliate abundance is coupled to maximum abundance of phytoplankton in the spring and early summer months. A high secondary maximum in the autumn is frequently found in eutrophic and mesotrophic lakes. Among dystrophic lakes that receive very high concentrations of allochthonous dissolved organic matter, microaggregates of particulate detrital material are abundant in the plankton.
Rotifers: Rotifers (Fig. 10) form an important group of soft-bodied invertebrates of the zooplankton. They exhibit a wide range of morphological variations and adaptations. In most, the body shape tends to be elongated and the regions of the head, trunk and foot usually are indistinguishable. The anterior end or corona of the rotifers is ciliated; in some species the periphery is also ciliated. The movement of cilia aids both in locomotion and movement of food particles towards the mouth. Most of the rotifers both sessile and planktonic are nonpredatory. Omnivorous feeding occurs by means of ciliary movement of living and detrital particulate organic matter into the mouth cavity. The maximum population of rotifers are associated with macrophytes; especially plants with finely divided leaves; densities commonly reach 25,000 per litre. The planktonic rotifers feed largely on sediment particles by the pulsating action of the coronal cilia. The size of food consumed varies, with most food particles taken being small, less than 12mm in diameter, although larger cells up to approximately 50 mm are seized, ruptured and their particulate parts ingested. Feeding behaviour of suspension-feeding rotifers is related to the type of particles, and food size, shape and
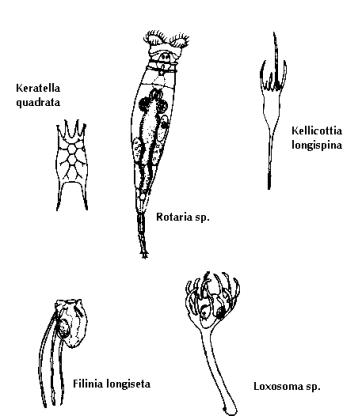
Fig 10: Rotifers
density. Food selectivity clearly occurs in some rotifers and is governed by several rejection mechanisms, either by screening particles or by rejection of ingested particles. Food limitations to the growth of rotifers vary widely. Threshold food concentrations for rotifers are high in relation to their small size in comparison to other zooplankton. In particular, large rotifer species have high threshold food concentrations that restrict them ecologically to nonpelagic environments.
A number of similarities exist among the reproductive processes of rotifers and cladocerans. During the main growing season, males are absent and reproduction occurs via parthenogenesis of diploid females. Under certain conditions of environmental stress, meiosis and sexual (mictic) reproduction occurs, leading to formation of mictic haploid cells. If fertilised, the diploid eggs (resting eggs) develop a heavy cell wall and become resistant to environmental extremes. The resting eggs commonly enter diapause, with hatching delayed for long periods; and upon return of favourable conditions, will develop into amictic diploid females. The survival and reproductive rate of rotifers are related strongly to the quality and abundance of food as well as temperature. These factors have major importance in determining seasonal fluctuations in the populations resulting from changes in the balance between increases by reproduction and decline from mortality.
Crustacean zooplankton: The crustacean arthropods are mostly fresh water aquatic, but some are marine. Respiration is accomplished through the body surface or gills. The body is differentiated into separate regions; but the tendency is toward fusion of the abdominal and thoracic segments. In fresh water, truly planktonic crustacea are dominated by cladocerans and copepoda. Most of the Ostracoda are benthic.
A) Cladocera: The suborder Cladocera includes mainly microzooplankton. With the exception of two species, nearly all cladocerans range in size from 0.2 to 3.0 mm. Light sensitive organs usually consist of a large compound eye and a smaller ocelleus. The second antennae are large swimming appendages and constitute the primary organs of locomotion. The mouthparts consist of (1) large chitinized mandibles to grind food particles, (2) a pair of maxilulles used to push food between the mandibles and (3) a median labrum that covers other mouthparts. Cladocerans have five pairs of legs attached to the ventral part of the thorax. The legs are flattened and bear numerous hairs and long setae. Complex movements of these legs create a current of water through the valves, which oxygenates the body surface and forces a stream of food particles anteriorly. The food particles filtered by the setae collect between the bases of legs and are impelled forward toward the mouth to be mixed there with oral secretions. Reproduction in the cladocerans is parthenogenetic during a greater part of the year. Until interrupted by sexual reproduction, females produce eggs that develop into more parthenogenetic females. The number of eggs produced per clutch varies from 2 in Chydoridae to as many as 40 in the large daphnidae. The eggs are deposited into a brood chamber or pouch, a cavity dorsal to the body bounded by the valves of the carapace. The eggs develop in the brood pouch and hatch as a small form of the parent. (Thus there are no free-living larval forms among Cladocerans).
B) Copepoda: The free-living copepods of this order of the class crustacea are separable into three distinct groups: three suborders Calanoida, Cyclopoida and Harpacticoida. The body consists of the anterior metasome (cephalothorax) which are divided into the head region, bearing five pairs of appendages, representing antennae and mouth parts, and the thorax with six pairs of swimming legs. The posterior urosome consists of abdominal segments, the first of which is modified in females as the genital segment. The three suborders of free-living copepods can be distinguished by the general structure of the first antennae, urosome and the fifth leg. The mouth parts of harpacticoids are adapted for seizing and scraping particles from the sediments and macrovegetation. Feeding is raptorial i.e., plant or animal food particles are seized by the mouth parts and brought to the mouth. The maxilulles hold and pierce the prey and force particles between the mandibles. Some particles are swallowed intact and differentially digested. Diatoms tend to be digested, while some green algae, if they are not ruptured, pass through the gut undigested. Reproduction in free living copepods is similar in spite of widely varying species differences in sexual behaviour and periodicity of breeding. Some species reproduce throughout the year, others only briefly at specific times of the year. Copepod copulation occurs by a male briefly clasping the female with transfer of spermatophores to the ventral side of the female genital segment. The female in 1 or 2 egg sacs carries fertilised eggs. The number of eggs in the cyclopoid copepods, carried in two lateral egg sacs is variable. Copepod eggs hatch into small free living larvae termed nauplii, and then develop by molting through a number of subsequent larval stages.
FISH COMMUNITY
Fish
are very diverse in the open-water community. They include omnivores eating a
range of foods from invertebrates to plants. The greatest number of freshwater
fish lives in the tropics. Feeding among fish, as among other animals, is the
process of acquisition of both energy and nutrients required for growth and
reproduction. The efficiency of this process is determined by both physiological
and environmental constraints. Nearly all fish use suction feeding to ingest
food, by which the fish creates a negative pressure in the buccal cavity and
draws into the mouth a stream of water carrying organisms. Intake can also be
accomplished by swimming with the mouth and opercules open. Size selection
of prey by fish is governed by energy return obtained per unit of energy
expended in foraging and by the abundance of prey. When prey are abundant, only
larger prey are consumed; as prey abundance decreases, small prey are henconsumed.
When prey density is high and size is small, fish commonly shift to filtering
particles from the water by entrapment on mucus or by mechanical sieving. The
filtration apparatus are commonly gill rakers.
Vision is critical to the location of prey among many fish and has led to many behavioural adaptations among prey organisms to seek both temporal and spatial refuges from the visual feeding of fish. The success of visual predation varies greatly with prey size, shape, translucency, colour as well as water quality, prey behaviour and other factors. Many fish adapt to a wide variety of fish sources and often switch from one food source to another as environmental and food supply conditions change dietary quality and abundance. The species flexibility to switch and to take advantage of the most profitable food source at any particular time, termed trophic adaptability, makes it very difficult to group fish into specialist or generalist fish. Specialist fish feed on a restricted diet, whereas generalist fish feed on a broad spectrum of prey species or detrital organic matter. Opportunist fish switch from one food source to another as food populations fluctuate and as their life stages mature. For example, larval fish utilise endogenous sources of nutrient in the yolk sac stages and shift to exogenous mouth ingestion of algae initially as the yolk sac is absorbed and to zooplankton, the latter being selected individually, in the later stages of larval feeding. Mortality among larval fish is very high and some has been attributed to starvation.
A number of fish families are predominately herbivorous on algae and vascular aquatic plants. All herbivorous fish ingest some animal food, which may supplement the largely plant diet. Detritivory is common among fish. The diet of adult detritivorous fish consists mainly of decomposed organic matter; living organisms associated with the detritus are also consumed, either actively or fortuitously. Early larvae of detritivores are zooplanktivorous and switch to dead organic matter as they mature.
|
Phosphorous cycle in the plankton: Phosphorous enters from the catchment or is released from the sediment. It leaves when washed through the outflows or permanently incorporated into sediments as detritus or precipitates. During the planktonic cycle, there are at least two main means of regeneration of inorganic phosphate, zooplankton and microorganisms. When nitrogen and phosphorous are released by zooplanktons they are rapidly taken by phytoplankton. About 12.5% of the phosphorous in the plankton is regenerated daily by excretion. Phosphate is taken in very rapidly by the smallest organisms and only slowly acquired by larger phytoplanktons through mineralization by microbial complexes and release of organic and inorganic phosphates. Phosphorous is thus rapidly cycled and reused by the smaller phytoplankton and bacteria. Organic phosphorous compounds may not always be available directly to the phytoplankton but should be metabolised by bacteria other than those, which produce and secrete it. Zooplankton also cycle phosphorous, at a longer turnover time. |
Diversity of feeding is great among bottom (benthic) foraging fish. Most species shift among prey types as availabilities change, but certain behavioural adaptations often predominate. Processes include (a) ingesting sediment deposits, (b) scraping periphyton off rock surfaces, (c) grasping invertebrates from surfaces and (d) crushing molluscs.
|
Nitrogen cycle in the plankton: The behaviour of nitrogen in the plankton is as complex as that of phosphorous. There are four main dissolved inorganic pools, ammonium, nitrite, nitrate and molecular nitrogen. Only certain blue-green algae and photosynthetic and heterotrophic bacteria can use molecular nitrogen. Blue green algae having heterocysts can fix nitrogen. The enzyme nitrogenase is inhibited by the presence of oxygen and lacks a part of the photosynthetic apparatus called as heterocyst, responsible for oxygen release. Blue green and other algae and bacteria secrete nitrogenous organic compounds in water, where they form part of a pool of many different dissolved organic nitrogen compounds. Excretion of urea by fish and amino acids and urea by some zooplanktons also supply nitrogen. The vitamins required by the phytoplankton species are all dissolved organic nitrogen compounds and are rapidly used once they are formed. Dissolved organic nitrogen compounds although not solely responsible may also act as chelators. Chelators reversibly combine with metal ions in equilibria between free metal ions and soluble ion-chelate complexes. As ions such as Fe 3+, Mn 3+ and Mo 3+ are removed from the complexes by phytoplankton, the equilibrium tends to replace them. Such ions are readily precipitated inorganically as hydroxides or carbonates and retention in soluble chelator complexes ensures a steady supply for the phytoplankton. The peptides secreted by blue green algae act as chelators. Algae: Some of the major ones are as follows - Green algae: All algae contain the green pigment chlorophyll, which is not masked by any other pigment or structure. Some green algae are mobile, some form colonies, while others are single cells. A few form long filaments of cells. A specialised group are the desmids, which come in many shapes and sizes but are typical of the soft waters. Blue green algae: In this type of algae, chlorophyll is present free in the cell and not in specialised structures called as chloroplasts. Like bacteria, they do not have a definite nucleus. They are a significant component of the phytoplankton in lakes with high concentrations of nutrients, particularly in warm water and in the tropics. Diatoms: The cells of this group are encased in rigid cell wall with high silica content and decorated with geometric patterns characteristic of each species. Dinoflagellates: Majority of this group live in the sea but Ceratium, a distinctive species, is found in freshwater. The cells have sculptured cell walls and two long flagellae, characteristic of the species, which helps them migrate through the water.
|
FRESHWATER BIRDS:
Freshwater is used by water birds, which feed on aquatic organisms, and other birds, which frequent the varied vegetation of water-land interface.

|

|

|

|
| Black Ibis | Cattle Egret | Coot | Common Sandpiper |
|
|

|

|

|

|
| Cormorant | Duck | Green Shank | Jacana | Painted Stork |

|

|

|
| Little Grebe | Little Egret | Little Ringed Plover |

|

|

|
 |
| Pond Heron | Small Blue Kingfisher | Stilts | Spotted Billed Pelican |
Habitat factors:
The main habitat factor involves the consideration of the process of succession by which vegetation and mineral soil encroach on freshwater areas, converting them into terrestrial habitats. The hydrosere describes gradients, over time from open water to terrestrial habitats. In fertile (eutrophic) situations, the gradient goes from open water with only submerged aquatic vegetation through deep and shallow marshes, with herbaceous emergent vegetation standing in water. In less fertile and more acidic situations, the (oligotrophic/dystrophic) gradient passes from open water often with rocky shores through water areas, with some emergent vegetation to wet areas with some standing water and shrubby as well as herbaceous vegetation. These processes are slow, progressing over several hundred years for completion, unless interrupted by changes in the water level.
Wetland Users:
Many birds characteristic of wetlands nest some distance away on dry land, often in a plant community in no way associated with wetlands. Others nest in vegetation above or adjoining the wetlands. Various groups lead their young from the nest, before these can fly, to rear them on open water in marshes or on shores. Nevertheless many birds may be assigned to particular stages of one or more gradients, though some tolerant species occur in nearly all wetland habitats.
Open water: Several water birds spend most of their time foraging and resting on the open water, but these do not nest over water as a rule. This is a heterogeneous group linked by their dependence on deeper open water for feeding.
Deep and shallow marshes: Birds that build floating nests anchored to floating aquatic vegetation seldom go ashore for any purpose. These along with other birds that nest in emergent vegetation comprise the marsh-bird community.
Wet pastures or meadow: Seasonally flooded wet areas, characterised by grasses, sedges and poorly drained soils are used by various species of birds. These habitats are generally associated with or altered by human activities.
Waterfowl Ecology and distribution:
Some of the major factors influencing the use of inland habitats by waterfowl include fertility, climate, shoreline gradient and water/land interspersion, and the disturbance of birds and their habitats.
Fertility: Insufficient nutrients in the soil and water limit the growth of vegetation and aquatic invertebrates used by waterfowl. Excessive salinity or acidity or other chemical factors can also inhibit growth of plants and animals even in the presence of adequate nutrients.
Climate: Factors include water level variability and its duration, and incidence of unfavourable rainfall/ temperature combinations during the hatching period.
Shoreline gradient and water/land intersection: A wetland with islands and a highly indented shoreline provides isolated breeding spaces for more ducks than would a similar area with some regular shoreline. The interspersion and combination of vegetation determine the use of the area by waterfowl.
Disturbance of birds and their habitat: Birds and their habitat are disturbed due to anthropogenic activities including motorboats, inflow of sewage and industrial effluents. Wetland and water birds make use of a variety of conditions, from dry zones and meadows bordering lakes to open water zones. On the basis of their size, the availability of food and suitable foraging conditions, different birds can occupy different parts of the lake. There are five major groups of water birds found based on the wetland zones they frequent. These are: (i) Open water birds, (ii) Waders and shoreline birds, (iii) Meadow and grassland birds, (iv) Birds of reed bed and other vegetation, and (v) Birds of open air space above wetlands. Ducks, geese, grebes, cormorants, kingfishers, terns, gulls and pelicans represent the open water birds. Stilt, greenshank, sandpipers, storks, ibises, spoonbill, herons and egrets tend to frequent shallow waters. Rails, bitterns, coots, jacanas, moorhens, snipe, painted snipe, etc. are found in vegetated portions of the wetland.