
LIMNOLOGY
LAKE
ZONES
A typical lake has three distinct zones (limnetic, littoral and the benthic zone; Fig. 11) of biological communities linked to its physical structure. The littoral zone is the near shore area where sunlight penetrates all the way to the sediment and allows aquatic plants (macrophytes) to grow. The 1 % light level defines the euphotic zone of the lake, which is the layer from the surface to the depth where light levels become too low for photosynthesis. In most lakes, sunlit euphotic zone occurs within epilimnion. In transparent lakes, photosynthesis may occur well below the thermocline, into the perennially cold hypolimnion. The higher plants in littoral zone, in addition to being a food source and a substrate for algae and invertebrates, provide habitat for fish and other organisms different from the open waterzones.
The limnetic (pelagic) zone is the open water area where light does not penetrate to the bottom. The third component of the lake habitat is benthic zone (the bottom of the lake), covered by fine layers of mud in which animals live. In the littoral zone, there is enough light for rooted plants to grow, but beyond this zone, there are no rooted plants as the water is too deep for light to reach them. The deepest part of the open water forms profoundal zone, but this is relevant only in extremely deep lakes.
In each community, green plants produce food upon which all aquatic organisms depend. Hence they are called as autotrophs. Animals, which feed on these plants, are called as herbivores and those that eat other animals are called carnivores. Omnivores have a mixed diet and detrivores feed on dead and decayed plant and animal material. All the feeding habits link to form the food-web.
Fish are usually the largest animals in the lake, feeding on most invertebrates; some eat other fish and very few are herbivores. There are more than 20,000 species of fish out of which 7000 live in fresh waters. Fish feed on plankton, plants and invertebrates and are in turn eaten by carnivores such as birds, otters and man. At each step, food is eaten to gain energy and nutrients. Most of the energy generated is used to support daily activity and is dissipated into the environment as heat. Only surplus energy is available for growth and reproduction, such that the energy available for the next link in the food chain is always less than that which entered the present trophic level. Hence, there cannot be more carnivores than herbivores.
Pelagic zone community:
Pelagic zone is the home of plankton and nekton. They are distinguished based on their swimming ability. Planktons are suspended in water whereas nektons are active swimmers. The phytoplankton consists of algae ranging in cells from 10 mm in diameter such as Chlorella to clusters of cells (Volvox). Several major groups of algae occur the world over, although the species vary. Majority of the planktonic animals (zooplankton) are herbivores feeding on algae, but there are also carnivores. The most important species are crustaceans of two major groups: cladocera (eg., Daphnia) and copepoda. They range in size from 0.2mm to about 4mm long, with exceptional species reaching 1-2 cm. These animals are an important link between the algae and much larger creatures such as birds and fish. Fish are the most obvious predators of zooplankton. Most fish start feeding on rotifers and move on to cladocerans.
A most unusual community called neuston lives at the air-water interface of the lakes. Numerous types of algae and bacteria attach themselves to the surface, taking advantage of the surface tension. The cladoceran, Scaphaloberis mucronata filter feeds on small particles while attached to the surface film. Typical pelagic fish include clupeids, which are an important link in the food chain of open water zones and a significant source of human food. Most birds feeding in this zone are fish eaters and either dive for their prey or catch it close to the surface. Birds (like geese and duck) are also attracted to these zones for refuge from terrestrial predators.
Benthic zone community:
The term benthic applies to flora and fauna living at the bottom and those species that live by burrowing in it. Benthic organisms can live in the substrate (in mud and sand), move on the substrate surface, grow attached to the surface or move freely in the bottom. Benthic primary producers include cyanobacteria, all higher taxa of eukaryotic algae and flowering plants. Algae attached to a substrate are called as benthic, to distinguish them from phytoplankton, which live floating in water. In some shallow lakes, benthic algae may be an important source of food but most benthic animals beyond the littoral zone derive their food from detritus raining down from the open water.
Much of the bottom beyond the littoral zone will be covered with mud whose particle size and organic content depends on conditions specific to a lake. The larger inhabitants of this zone are mostly worms, larvae of chironomid flies, and molluscs. In addition there are numerous smaller animals like nematode worms and ostracods. Chironomids are very characteristic lakeside inhabitants, typical of many lake insects, with larvae living and growing in the water. The larvae are essential food for many fish and ducks. The phantom midges, Chaobrus spp, also have aquatic larvae whose adults emerge out from the water. They are interesting as they provide a link between benthos and pelagic zones. During the day, they rest in the substrate as members of the benthos and at night, they raise upwards in the water becoming pelagic to feed. They are voracious carnivores whose predation has a marked effect on other zooplankton. Many of the worms found in fresh waters have haemoglobin in the blood, which help trap oxygen, an advantage, as the benthic community suffers from lack of oxygen.
Molluscs are important components of the benthic fauna. The molluscs that are adapted to the benthic life are the burrowing bivalves, such as freshwater mussel. Mussels are important filter feeders and are usually the largest members of the lake community to feed directly on the smallest algae. The animals, which live in the deepest part of the lake water, are able to extract oxygen even from water with less oxygen content. At about 20-25% saturation, their respiration rates decrease, but they can survive periods of low oxygen concentration, although their feeding and growth are slower. Benthic animals have to tolerate long periods in summer, when not only the sediment but also the water lying on top is deoxygenated. Unlike pelagic fish, benthic fish cannot find their prey by sight, and many feel and taste for it with barbells (whiskers) around their mouth. Carps are typical benthic fish, which feed on whatever is available (molluscs, crustaceans, insects and plants), consuming more at higher temperatures.
Littoral zone community:
The main groups of benthic invertebrates are also found in the littoral zone, but are often represented by different species. The inshore area is a more complex habitat, mainly due to the presence of macrophytes and supports many animal species. The only substrate for animals in this zone is rocks and the only food is algae either present in the water or attached to the rocks, and the detritus that is lodged among the stones. The advantage in this zone is that oxygen is never likely to be limited as water is constantly moving, dissolving more oxygen from the air.
Snails with a heavy shell and strong muscular foot are common, particularly if calcium content is high. Sponges encrust the rocks and leeches are common. Worms and freshwater shrimps are among the organisms that escape the waves by hiding. The common worms, larvae of caddis flies and chironomids, either attach themselves under the stones or burrow among the pebbles.
In sheltered places where sediments can accumulate, plants grow and make a tremendous difference to the number and type of animals found in this marginal area. Large plants in a lake are referred to as macrophytes, distinct from the other major plants, algae, most of which are microscopic. Most macrophytes need to be rooted in the bottom silt and therefore grow in relatively shallow water. Those at the lake edge normally grow tall and stick out of the water, referred to as emergent macrophytes. On gentle slopes, the most obvious macrophytes are tall aquatic grasses, notably reeds (Phragmites). The flat floating leaves of plants such as water lilies (Nymphaea) and pond weeds (Potamogeton) are rooted but have long stems. Further out in the lake are wholly submerged plants, some of which are rooted to the bottom, such as water milfol (Myriophyllum) and hornwort (Ceratophyllum) that float freely in the water. Among the plants that float on the surface in sheltered places, the smallest are duckweeds (Lemna), and the largest include water hyacinth (Eichhornia) and floating fern (Salvinia).
The gradation of plants from land into water represents a transition from one environment to another and is known as zone of succession (hydrosere). As the emergent macrophytes die each year, their remains accumulate and trap debris and silt. Emergent macrophytes are similar to terrestrial plants except that they are always rooted in deoxygenated mud rather than well-aerated soil. Like other parts of the plant, the root cells need oxygen and if this is not obtainable from their immediate surroundings, it must be transferred down the stem from those parts of the plant, which are out in the air. For this purpose, roots, leaves and stems of macrophytes contain large air-filled spaces along which oxygen can travel to relieve the shortage (of oxygen) in the roots. Wholly submerged plants cannot obtain oxygen from the air and must extract all they need from water. They have very thin leaves and much of the oxygen released during photosynthesis goes to alleviate the shortage in the roots.
Animals in the littoral zone mostly live on the surface of leaves or stems or burrow among the plant roots. The littoral zone contains a complex mixture of plants, animals and microorganisms. Diatoms and blue-green algae are common components and the latter secretes a gelatinous layer, which attracts and holds other organisms. Pond snails are a conspicuous group of littoral zone animals along with many species of crustacea (shrimps and crabs) living among the plants. Littoral zone provides habitat for insects like dragonflies (Odonata), mayflies (Ephemeroptera), stoneflies (Plecoptera), caddis flies (Trichoptera) and midges (Diptera).
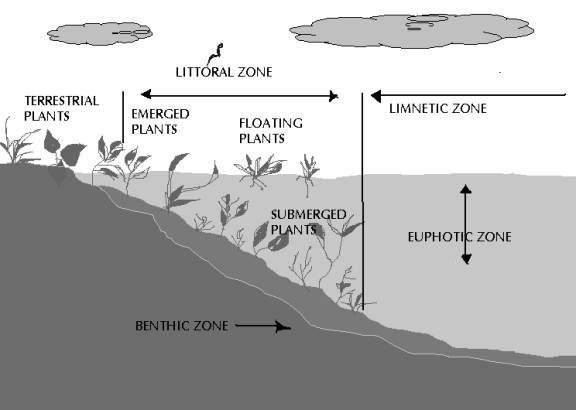
Fig 11: Lake Zones
COUPLING OF HABITATS:
Benthic and pelagic habitats are not completely separate from one another as there are numerous connections between their components. Deepwater fauna is very dependant on the production in the euphotic zone. Animals can greatly accelerate the vertical transport of organic matter. For example, the faecas of zooplankton, especially copepod faecal pellets are an important source of food for the benthos. These faecal pellets contain a large amount of partially digested algae. Due to their large size, they sink to the bottom before the organic matter is decomposed.
Smaller lakes have more connections between the littoral and pelagic zones. Phytoplankton carried to the littoral zone provides food for mussels and cladocera. Some animals are littoral in one part of their life cycle and benthic in another such as some cyclopoid copepods, which are planktonic as juveniles and adults, but spend a part of their development as a resting stage in the sediments. During their development, fish change their habitat according to the location of the food. As juveniles, many cyprinids for example, eat plankton and therefore stay in the pelagic zone. When they become larger, they shift to feeding on the bottom during the day and move into the littoral regions of the lake at night. Some planktivorous fish spend the day near shore and move into open water at night to feed on pelagic plankton.
PRODUCTIVITY
Productivity is the rate of production for a given group of organisms, essentially representing the net growth rate of organisms. Lakes seldom suffer from shortage of water but are often unproductive due to lack of nutrients necessary for plant growth and reproduction.
Plants comprise the basic food material, directly or indirectly, for the whole lake ecosystem. Their abundance can therefore indicate the lake’s productivity. Productivity is measured as the new carbon collected from the air and fixed as organic compounds by photosynthesis, so that it can be added to the total food supply in the lake. It is expressed as grams of carbon fixed per cubic metre (g C/m3) per unit of time. The amount of small floating plants or phytoplankton indicates productivity, measured as the total amount of chlorophyll present. The amount of chlorophyll-a per cubic metre is an index of phytoplankton abundance and potential productivity.
Efficiency of carbon transfer within lake ecosystems is dependant on two factors: a) quantity of biomass created at primary producer level and b) composition of species, which determines efficiency of grazing and quality and quantity of fish (representing the highest trophic level in the system). After death, organisms at primary, secondary and tertiary levels decay and recycle nutrients.
Increased
supply of nutrients due to anthropogenic activities (sewage, industrial
effluents, fertiliser run-off from agricultural fields) changes the lake
ecosystem causing eutrophication. Lakes with low supply of nutrients are called
oligotrophic, while those with high nutrients are called eutrophic. Continuous
eutrophication results from autotrophic production of internal organic matter by
primary producers (photosynthetic plants and algae) from nutrients available in
the lake, derived either from external sources (anthropogenic activity in the
catchment) or internal recycling (decay of organic matter and dissolution from
bottom sediments). In many shallow lakes, eutrophication may be manifested in
macrophytic growth rather than phytoplankton. Efficient utilisation of nutrients
depends on several factors, which define the growth condition, and hence the
resultant biomass production at the primary producer level. Grazing of
phytoplankton by zooplankton (secondary producers) and predation by fish
(tertiary consumers) represent carbon transfer system of the lake.
Eutrophication can be highly detrimental to the quality of lake water and may limit its use. The detrimental effects may result from inefficient use of phytoplankton biomass that in turn is derived from high nutrient availability. This is mainly due to changes in dominance of algal species that are not consumed or inefficiently consumed by zooplanktons.
Lakes are classified based on productivity as follows:
· Eutrophic lakes: These display high concentration of nutrients, high biomass productivity and low transparency. Oxygen concentrations can get very low (as low as 1 mg/L) in the hypolimnion during summer.
· Hypereutrophic lakes: These are lakes at the extreme end of eutrophication with very high concentration of nutrients and associated biomass production. Anoxia or complete loss of oxygen takes place in the hypolimnion during summer.
|
Changes, which occur in the lake due to addition of nutrients (on account of human activities), are called as eutrophication. The additional nutrients (primarily nitrogen and phosphorous) most usually come from sewage, industrial effluents or agricultural fertilisers, or all the three. Bacteria that use up the oxygen in the water first decompose raw sewage. Sewage always contains a higher concentration of phosphates than nitrogen due to increased use of phosphate containing detergents. In unpolluted waters, growth of algae is inhibited by the lack of phosphorous but the addition of these nutrients (through domestic sewage, other effluents etc.) increases algal growth in the receiving waters. While algae produce oxygen during the day due to photosynthesis, they use it up in the night. Thus, dense algal blooms cause suffocation of fish and other aquatic organisms. Increased algal production due to eutrophication initiates other changes in the aquatic ecosystem. |
Detritus -Dead organic matter and detrital dynamic structure:
Living biota constitute only a small portion of the organic matter of living systems, even in the most productive of freshwater habitats. Much of the organic matter in aquatic ecosystems is non-living and is called as detritus. The functioning of aquatic ecosystems depends on the cycling of organic carbon between living and non-living components. Detritus consists non-living organic matter in both dissolved and particulate forms. In aquatic ecosystems, organic compounds depend on dissolved organic carbon compounds (DOC) and particulate organic compounds (POC), the ratio usually being 6:1.
Organic matter originates from photosynthesis within the lake or the drainage basin. Organic loadings in particulate form are very rare and all organic carbon is transported in the dissolved form, which leads to stratified sedimentation, metabolised or exported. Very little of either the particulate or dissolved organic matter emanating from new photosynthetic production is consumed by animals or enters the consumer food chain, but enters dissolved and particulate detritus. These microbial and chemical fluxes are essential to the maintenance of nutrient recycling and metabolic stability of the ecosystem.
The amount of detrital organic matter is quite large and is chemically more recalcitrant to biological degradation than living organic matter. As a result, metabolism of detritus is slow but the quantity is large and much greater than that resulting from living organic matter ingested by animals. This slow degradation provides an inherent ecosystem stability that dampens brief volatile fluctuations of higher trophic-level populations, as they respond to these rapidly oscillating factors governing their growth and reproduction.
Seasonal Changes
in the lake community:
A lake’s biological characteristics are determined largely by physical characteristics of the water column. Important physical characteristics include temperature, light transparency, as well as the total abundance of inorganic nutrients (largely a watershed characteristic). In addition, preceding populations influence successive populations by the assimilation of critical nutrients. As physical, chemical and biological conditions in the lake change over time, some species will be effectively eliminated from a lake those which cannot tolerate new conditions. Other species will be out-competed by organisms that are better adapted to the new environment. Some species flourish for a period of time and then give way to other species more compatible with changed conditions, such as warmer water, more daylight or lower concentrations of nitrogen and phosphorous. Short-lived plankton communities are characterised by these seasonal fluctuations; longer-living organisms such as fish, must be tolerant to different lake conditions throughout the year.
Phytoplankton community: Depending on the species, algae divide to form new individuals, two to ten times per month. It is possible for populations of algae in lakes to exhibit a succession of species during the course of one year. During stratification, the replacement of one dominant species by another may be determined by chemical factors or grazing effect of animals. Some species are present all the time but fluctuate in numbers, but many others show a clear seasonality and disappear for the remainder of the year. The time of appearance each year is characteristic to different species. In general, there is a tendency for diatoms to dominate in spring, followed by green algae and blue green algae later in the year. Generalisations are difficult to make because of the great variability observed among phytoplanktonic numbers and biomass from lake to lake. Several patterns are reasonably consistent, however:
a) The seasonal periodicity of phytoplanktonic biomass is reasonably constant from year to year, if the freshwater ecosystem is not disturbed by outside influences, such as human modifications of the watershed, nutrient loading, etc.
b) The extent of change in phytoplanktonic numbers and biomass through the seasons is usually very high, a thousand fold in temperate waters. Whereas seasonal variations in tropical waters are much lower, often as little as fivefold.
c) Maxima and Minima of numbers and biomass of phytoplankton often are out of phase with rates of primary production. Primary productivity usually follows closely the annual cycle of incident solar radiation in temperate lakes.
Insects:
Lake insects respond to seasonal cycles. Adults of different species emerge at
different times of the year, such that there is seasonal succession of species.
As the insects grow larger, they provide food for fish; the fact that each prey
species reaches a maximum size at different times means that there is always
food available for the predators that switch progressively from one species to
the other.
Zooplankton: These populations also exhibit seasonal changes in their dominant species. Rotifers can increase their populations rapidly, and in strongly seasonal lakes are first to respond to increased algal populations in spring. Seasonal variations in flagellate and ciliate abundance are variable. Cryptomonad and other microflagellates are commonly numerically dominant across a spectrum of oligotrophic through hypereutrophic lakes. Generally ciliate abundance is coupled to maximum abundance of phytoplankton in the spring and early summer months. A secondary maximum in the autumn is frequently found in eutrophic and mesotrophic lakes.
Fish: Fish have a clearly defined breeding season and there is one period of the year when larval fish are abundant in the lake. In most lakes, where there is more than one species, their breeding may be separated in time so that the larvae of one species do not compete with those of another for the available food. In tropical lakes, some fish breed all through the year while others confine to breeding only during periods of flooding when they can breed in the shallow inshore areas or in the rivers which flow into the lake.
The
Energy Relationships of Freshwater Ecosystems
There
are four main constituents of the environment alongside abiotic factors that
form the freshwater ecosystem, which are as follows.
a.
Elements
and Compounds of the
ecosystem that are absorbed by organisms, required for food or respiration, many
of which are also required by plants.
b.
Plants,
which are autotrophic by nature, meaning they synthesize food by harnessing
energy from the sun through photosynthesis. These plants (and some bacteria) are
the primary producers, as they produce (and introduce) new energy into the
ecosystem.
c.
Consumers,
which are the organisms that feed on plant organic material. These may be
primary consumers, which feed on plant material or secondary consumers, which
feed on primary consumers.
d.
Decomposers
attain their energy by breaking down dead organic material (detritus), releasing
critical elements and compounds, which in turn are required by plants.
Biological communities within lakes may be organised conceptually into food chains and food web. The simplest illustration of the organisation of the organisms within an ecosystem is the ecological pyramid (food chain). The broad base of primary producers mainly supports overlying layers of herbivores, planktivores and less number of carnivores (predators). Food chains that allow a diversity of species to survive are divided into trophic levels, with plants providing the first trophic level, as they are the primary producers of most food chains. In almost all fresh water ecosystems, animals will be present, and form part of the many grazing food chains in the area. Other organisms leach the energy from dead organic matter forming detritus food chains. Such relationships allow the free flow of organic energy to be passed along from species to species, and provide an environment where food is available for them to survive. Food webs (Figs. 12 to 14) may be described in terms of both energy and nutrient (carbon, nitrogen or phosphorous) flows. The process typically begins with sunlight driven photosynthesis and respiration. Green plants capture energy from sunlight and convert it into living organic plant tissue. Lake photosynthesis includes algae and macrophytes, which together are the primary producers as they create organic material required by other organisms for nutrients and energy. Other fish feed on primary consumers and are called secondary consumers. They constitute the third trophic level. Still larger consumers such as large fish and man are tertiary consumers (fourth trophic level). Hence, energy and nutrients originating from the photosynthetic production of biomass and energy cascade through the food web. Respiration (the oxidation of organic material) releases energy that was originally captured from sunlight through photosynthesis. Decomposers are sinks for plant and animal wastes, but also recycle nutrients for photosynthesis. The amount of dead material in a lake far exceeds the living material. Detritus is the organic fraction of the dead material in a lake and is in the form of small fragments of plants, animals and microorganisms.
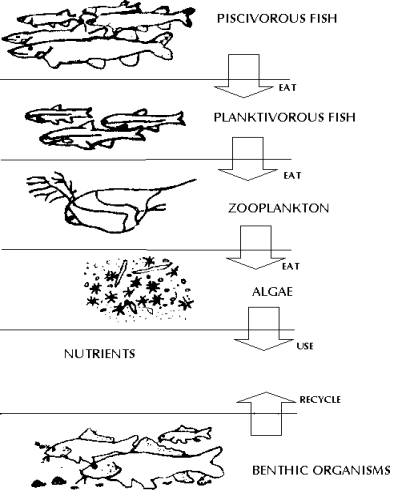
Fig
12:
Food Web
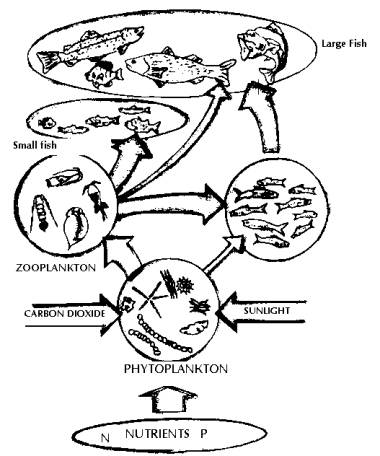
Fig
13:
Food Web in a Typical Lake
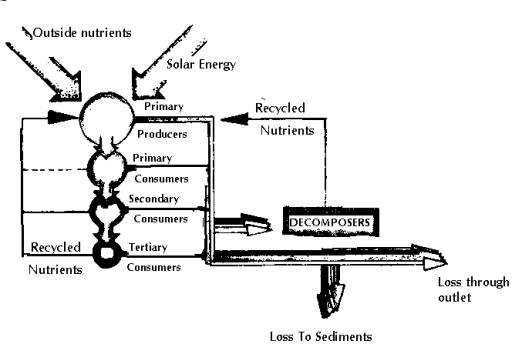
Fig
14: A Simplified Food and Energy Web in a Lake
Figure
15 indicates an ecological pyramid where energy is passed along from autotrophic
organisms to carnivores at the top of the chain. Those at the bottom of the food
chain are usually the smallest (not always) and are almost inevitably the
largest in number. Those which feed off these primary producers are less in
number, usually because they are larger and require more than one primary
producer as a means of food in order to survive. This situation continues to the
top of the chain, where few secondary consumers are eaten by an even smaller
amount of tertiary consumers.
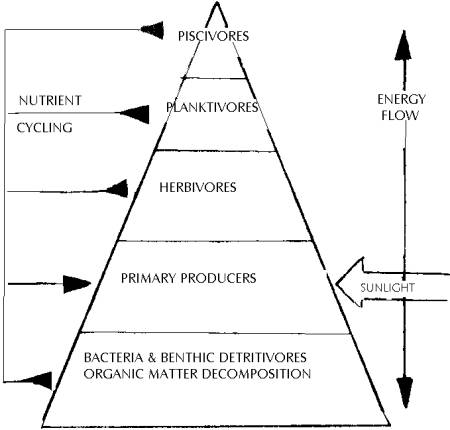
Fig 15: The Ecological Pyramid
Oxygen released during photosynthesis adds to oxygen supplied by the atmosphere. In water layers where photosynthetic rates are high, such as during algal blooms, the water may become supersaturated, the saturation depending on the temperature of the water. Cold water can hold more oxygen than warm water. During periods of stratification, the only potential source of oxygen to the deeper lake is photosynthesis. The whole interaction of photosynthesis and respiration by plants, animals and microorganisms represents the food web. Food webs are usually complex and in many lake ecosystems, hundreds of different species can be involved. As the available energy decreases at each trophic level, a large base of plants (primary producers) is necessary to support few fish. These plants die and decompose or are eaten by primary consumers (second trophic level). The second trophic level involves zooplankton grazing on algae and eating fish, and invertebrates that feed on periphyton.
The most important fact to be understood here is that no matter what species occupies an area, chances are they will require another species to be in abundance in the area for feeding. The population of a particular species will depend on density dependant and density independent factors. Therefore the abundance of food in the ecosystem (a density independent factor) comes into play. The prey of a particular species will also require the existence of a food source, and other conditions conducive for them to survive.
Organisms within a food chain that can be assigned to the same position are said to occupy the same trophic level. Because ecological efficiency is always low, there is a rapid decrease in the productivity of subsequent trophic levels. Ecological efficiency refers to the ratio of predator productivity to prey production. This is the measure of energy passed from one link of the food chain to the next. Ecological efficiencies range from 5 to 20%, i.e., 80 – 95 % of the energy is lost at each transfer in the food chain. Because of these losses, energy must flow in one direction in the ecosystem. Trophic pyramids allow one to visualise the decreasing production with increasing height of the trophic level. The blocks that represent the production rates become smaller as one goes up to the higher trophic levels, giving rise to the appearance of a pyramid.
CYCLING OF MATTER:
Transfer
of energy in ecosystems is always coupled with transfer of matter. The most
important difference is that matter can circulate in an ecosystem whereas energy
can only flow through the ecosystem. The biochemical cycles occurring in the
lake are not closed. Lakes receive substances from the watershed and atmosphere;
they export substances via outflow and atmosphere and bury substances in the
sediments. Depending on the balance of these processes, water passing through
the lake can be either enriched or diminished of particular compounds.
Modifications of the water in the watershed:
Before entering the lake, water can be chemically altered, while passing through the lake watershed. The geology of the watershed and the chemical composition of the precipitation are extremely important. On its way to the lake, water is modified mainly by rock weathering. This leads to an enrichment of bivalent cations, especially calcium. The increased content of calcium permits uptake of carbon dioxide from the atmosphere. Depending on the pH, CO2 is converted into bicarbonate and carbonate. Recycled matter from dead organisms also dominates nutrient loading in fresh waters. There are fundamental differences in the geological origin of different nutrients, such as nitrogen and phosphorous. Nowadays, much of the nitrogen and phosphorous entering the lakes is derived from soils. Human activities can greatly accelerate the release of nutrients into the lakes and streams. Non-point source pollution due to agricultural activities tend to increase the N:P ratio in the water bodies, while point source pollution due to domestic sewage (being rich in P), lowers the ratio of N:P.
Modifications of the water within lakes:
Water from the watershed enters a lake from surface and through groundwater inflows. The retention of a particular element may be calculated as the difference between the total amount of the element entering the lake via the surface and groundwater inflows minus the total amount leaving the lake provided the exchange with the atmosphere is negligible. If the retention capability of the sediment exceeds its threshold value, then the sediment releases substances instead of trapping them, causing a negative retention. However, negative retention is possible only for a short time, for those substances that have been previously deposited can be released from the sediments. In the long run, the sediments act as a sink for substances in the lake.
Balances are more complicated for elements that are exchanged with the atmosphere, like carbon and nitrogen. Exchange of gases with the atmosphere tends to reduce the degree of under-saturation and super-saturation in the surface waters. The net balance of exchange depends on biological processes in the lake. A lake, heavily loaded with organic matter and high rates of respiration by heterotrophic bacteria may have a net export of CO2. The total balance for nitrogen in the lake depends on the balance between nitrogen fixation and denitrification. Nitrogen fixation lowers the concentration of nitrogen in water, allowing for an influx of this gas from atmosphere. Denitrification increases nitrogen in the water and may result in export from the surface water to the atmosphere.
Carbon cycle:
Dissolved Inorganic Carbon (DIC), Dissolved Organic Carbon (DOC) and Particulate Organic Carbon (POC) are the most important pools of carbon in aquatic ecosystems. DIC consists of carbon dioxide, bicarbonate, and carbonate and is indirectly affected by pH, and photosynthetic and respiratory activity of the aquatic organisms. The most important input of DIC is from atmospheric CO2 and the dominant abiotic output is release of dissolved carbon dioxide into the atmosphere. Respiration is the biological input and photosynthesis is the biological output. In addition to allochthonous inputs, the most important sources of DOC are secretion and excretion by organisms from all trophic levels and autolysis of detritus. Uptake by heterotrophic microorganisms especially bacteria is the most important source of DOC. Excretion products are rapidly utilised by bacteria. POC consists of carbon bound in organisms and detritus. Primary production is the source of POC. There is shift in the POC pool due to organism death, feeding, parasitism, etc. POC can be transformed into DIC by respiration and into DOC via secretion, excretion and autolysis. POC is lost from the pelagic region and imported into the benthic zone through sedimentation.
Nitrogen cycle:
The most abundant nitrogen component in lakes is dissolved elemental nitrogen; however, only few organisms can utilise this form. Nitrogen fixation requires the enzyme nitrogenase that is found only in prokaryotes. For autotrophic organisms that do not produce nitrogenase, dissolved nitrite, nitrate and ammonium are the most important inorganic nitrogen forms. These forms of nitrogen are carried into the lakes through surface water, ground water and precipitation. Autotrophic organisms can use all three forms. But the decomposition of N-rich organic compounds releases only ammonium. In contrast to vertebrates that excrete either urea or uric acid, zooplankton excrete ammonium. The nitrate respiration that takes place in anoxic conditions transforms nitrate to either ammonium (nitrification) or elemental nitrogen (denitrification). Nitrification, which takes place under aerobic conditions, transforms ammonium into nitrate (through the intermediate formation of nitrite). These three forms of nitrogen are taken up by phytoplankton, and hence have a minimum concentration at locations where photosynthetic activities are maximum. Dissolved organic nitrogen comes from the excretion of organisms and decomposition of detritus. This consists of polypeptides and other amino acids. Simple amino acids are taken by bacteria and are therefore present in low concentrations. Microorganisms also excrete peptidases that assist in the splitting of peptides into smaller units.
Phosphorous cycle:
Phosphorous occurs only as phosphates in ecosystems. It is bound to organic compounds by ester bonds between phosphoric acid derivatives and carbon chains. Orthophosphates is much less mobile than nitrogen in the sediments and precipitates easily as it combines with several cations to form highly insoluble products. The most important material sink of soluble reactive phosphorous (SRP) is uptake by algae and bacteria. The carbon heterotrophic bacteria can be P-autotrophic i.e., in taking up the inorganic phosphorous they compete with phytoplankton for soluble reactive phosphorous. Despite the fact that phosphorous is often the growth-limiting factor, algae and bacteria lose soluble reactive phosphorous and to a lesser degree, low molecular weight non reactive phosphorous. During the growth period, soluble reactive phosphorous in the epilimnion may be reduced to its limit of detection. Periods in which zooplankton grazing exceeds phytoplankton production can usually be recognised by an increase in SRP concentration. SRP and particulate phosphorous have complementary seasonal changes.
Phosphorous moves into sediments through sinking of organisms and adsorption on sinking clay minerals. The redox condition in the sediment-water interface is of critical importance in determining the fate of phosphorous in the sediments. Concentration of phosphorous is usually higher in sediments than open waters. According to the concentration gradient, there is always a diffusion of phosphorous into the open water. When the deep water and sediment-water boundary is oxidised, iron occurs as Fe 3+ form. Phosphorous that diffuses up from the sediments forms an insoluble complex with ferric hydroxide. If the ferric is reduced to ferrous, the complex dissolves and can go back into solution. Phosphorous accumulates in the anoxic hypolimnion, since it remains soluble there and does not precipitate back into sediments.
![]()
