 |
Two new species of Gomphonema (Bacillariophyceae)
from Doon Valley, Uttarakhand, India |
 |
Balasubramanian Karthick 1*, Rachna Nautiya1,2, J. Patrick Kociolek3 , T. V. Ramachandra4
1Biodiversity and Paleobiology Group, Agharkar Research Institute, G. G. Agarkar Road, Pune 411004,
Maharashtra, India
2 Zoology Department, Government Post Graduate College, Dakpathar Vikasnagar, Dehradun 248125,
Uttarakhand, India
3Museum of Natural History and Department of Ecology and Evolutionary Biology,University of Colorado
, Boulder 80309,
CO, USA
4Energy and Wetlands Research Group, Centre for Ecological Sciences, Indian Institute of Science,
Bangalore 560012, Karnataka, India
Citation: Balasubramanian Karthick, Rachna Nautiyal, J. Patrick Kociolek, T.V. Ramachandra. Two new species of Gomphonema (Bacillariophyceae)
from Doon Valley, Uttarakhand, India,Nova Hedwigia, Beiheft 144, p. 165–174 Article
Stuttgart, July 2015.
Discussion
Gomphonema juettnerii resembles Gomphonema pumilum var. elegans Reichardt et Lange-Bertalot
in general shape (Reichardt 1997). It differs, however by less radiate striae, ultrastructure
of interstriae trench and external areola structure; areolae of G. pumilum v. elegans show well
developed “C” shape across the valve (Figs 2, 3, Tafel I in Reichardt 1997), whereas G. juettnerii
shows “C”-shaped structure within the central area, and areolae become reduced to comma
shape towards the poles. The central area of G. pumilum v. elegans is formed by shortened striae
(Figs. 1-11, Tafel II in Reichardt 1997), which are absent in G. juettnerii. In LM, G. juettnerii.

Figs 23 – 25. Scanning electron micrographs of Gomphonema doonensis. 23. External view of whole valve
showing double rows of areolae across the valve and undulate raphe. 24. Internal view of the whole valve
showing striae pattern and central area. 25. External girdle view showing the areolae opening, apical pore
fields and girdle bands. Scale bar represents 1 μm.
resembles G. minutum Agardh in general valve outline. However, these species differ from
G. juettnerii by absence of the central area created by shortened striae. Furthermore, G. minutum
has striae composed of two rows of areolae (Fig. 91 in Kociolek & Kingston 1999). Striae pattern
and absence of stigmata in Gomphonema juettnerii differentiate it from G. sinestigma Reichardt,
Jüttner & Cox (Jüttner et al. 2004).
Gomphonema doonensis resembles some of the tropical species of Gomphonema, including
G. eurycephalus Spaulding & Kociolek (Spaulding & Kociolek 1998) described from Madagascar
and G. latistigmata Kociolek, Passy & Lowe (Passy et al. 1997) described from South
Africa. Gomphonema doonensis has narrower valves and the presence of shortened striae in the
central area and these features distinguish it from G. eurycephalus. Gomphonema doonensis dif
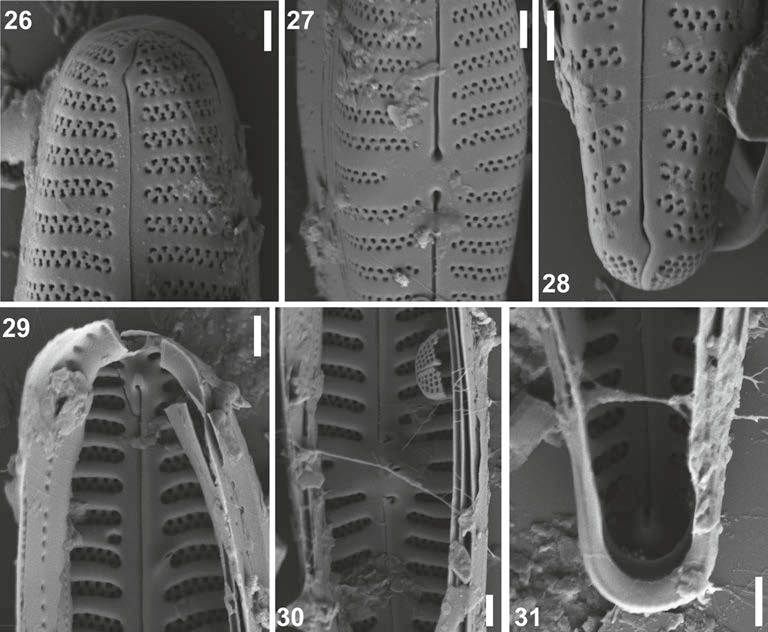
Figs 26 – 31. Scanning electron micrographs of Gomphonema doonensis. 26. External view of the headpole
with the external distal raphe end curving onto the valve mantle. 27. External view of the central area with
undulate raphe has dilated proximalends and round external stigmal opening. 28. External view of the
footpole showing distal raphe end and bilobed apical pore field. 29. Internal view of the headpole showing
striae found in deep troughs, separated by thick interstriae. 30. Internal view of the raised central area showing ecurved proximal raphe ends and a slit like internal stigmal opening. 31. Internal view of the footpole
showing the radiate striae and helictoglossae and pseudoseptum. Scale bar represents 1 μm.
fers from G. latistigmata in valve outline and length: the former is generally shorter and clavate,
with broadly rounded headpole.
There is considerable confusion as to whether presence of doubly punctate striae alone is both
necessary and sufficient to assign gomphonemoid diatoms to the genus Gomphoneis Cleve. Doubly
punctate striae were first identified by Hustedt (1942, 1945) in several new species of Gomphonema.
Hustedt noted (1942, p.121) that there were two features that diagnose Gomphoneis,
namely doubly-punctate striae and the presence of longitudinal lines. The latter character was not
mentioned by Dawson (1982), who documented doubly-punctate striae in the common diatom
Gomphonema olivaceum Hornemann using transmission and scanning electron microscopy. For
this reason, Dawson (1982) subsequently transferred G. olivaceum to Gomphoneis. Patrick & Reimer(1975, p.147) further complicated the situation by stating “This genus [Gomphoneis] is distinguished
by the double rows of decussating puncta alternating with the costae, and the narrow
septum near the margins of the valve.” Confusion over the taxonomy of this group was expressed by Lange-Bertalot (1980). In his identification of a large, doubly-punctate Gomphonema species
from Israel (as “Gomphoneis herculeana (Ehrenberg) Cleve”, a species known only from North
America!) and his argument was that doubly-punctate striae could not diagnose Gomphoneis.
Kociolek & Stoermer (1989) showed in the context of a formal phylogenetic analysis that Gomphoneis
is diagnosed by the presence of doubly-punctate striae and the presence of a marginal
lamina that gives the impression of the longitudinal lines in primitive members of the genus.
Furthermore, Kociolek & Stoermer (1989) documented that two groups may be recognized: the Elegans group (that is the name bearer for the genus, and whose species also either have no stigmata
or 4 stigmoids, as well as apical pore fields that are undifferentiated from the areolae) and
the Herculeana subgroup (more commonly referenced when discussions of the genus are made).
Krammer & Lange-Bertalot (1991) echoed Lange-Bertalot’s (1980) mistake when they suggested
that some species included the Elegans group of Gomphoneis found in Lake Baikal were Gomphonema
taxa, and made nomenclatural changes reflecting that opinion. See Kociolek et al.
(2014) about the diversity and high levels of endemism of Gomphoneis species in Lake Baikal.
More recently Reichardt (2007) argued on similar lines, while recognizing some gomphonemoid
species with doubly-punctate striae and assigning them all to Gomphonema, without considering
other features. To be clear, as Kociolek (1998) noted, we should refrain from thinking typologically about how species are included into groups (such as genera), and we have to recognize that there may be the loss or gain of evolutionary novelties. Formal phylogenetic analyses will guide these determinations, not a priori determinations made without such analyses. To date, a
morphology-based phylogeny of the freshwater gomphonemoid diatoms (Kociolek & Stoermer 1993a) suggests that a suite of features diagnose Gomphonema. These features include: differentiated
apical pore fields, a single, “typical” stigma (or in a few cases no stigma), no marginal
laminae and no axial plates, and with doubly-punctate striae with flaps occluding the areolae.
Diatoms described here from the Himalayas are placed firmly in the genus Gomphonema. We
are far past the time to be constrained by typological, non-evolutionary, approaches to taxonomy
and systematics.
|
|
T.V Ramachandra,
Centre for Sustainable Technologies, Indian Institute of Science,
Bangalore 560 012, India.
Tel: 91-080-23600985 / 2293 3099/ 2293 2506,
Fax: 91-080-23601428 /23600085 /2360685 (CES TVR).
Web: http://ces.iisc.ac.in/energy, http://ces.iisc.ac.in/foss
|
|