| Previous Session | Paper 1 | Paper 2 | Next Session |
SESSION-17
PAPER-1: Consequences of Catchment Processes and Climate Changes on the Ecological Status of Large Shallow Temperate Lakes
Tiina N õges 1,3 , Arvo Järvet 2 , Reet Laugaste 1 , Enn Loigu 4 , Ülle Leisk 4 , Ilmar Tõnno 1,3 , Peeter Nõges 1,3
CONTENTS-
Abstract
Introduction
Changes in Riverline Discharges
Changes in River Water Chemistry
Changes of Nutrient Loading
Consequences of the Changed Nutrient Loading on the Phytoplankton of Lakes
Consequences of the Changes of Global Climate on Water Level and the State of the Ecosystem
Acknowledgements
References
| Abstract | up | previous | next | last |
Synopsis: Watershed of L. Võrtsjärv (3 104 km 2 ) belongs to the basin of L. Peipsi (47 800 km 2 ). Regular investigation of these large, shallow, nonstratified lakes (L. Peipsi: area 3 555 km 2 , mean depth 7.1 m; L. Võrtsjärv: 270 km 2 , 2.8m) started in the 1960s. In the 1980s, the riverine nutrient discharge increased drastically while in the early 1990s a sharp decrease occurred, especially in the loadings of nitrogen. As nitrogen loading decreased faster than phosphorus loading, the N/P loading ratio decreased. There has been an increase in cyanobacterial dominance in the late 1960s and early 1970s, decline in the late 1970s and 1980s, and a new increase in the late 1990s in both lakes. Since the beginning of the 1990s, the biomass of N 2 -fixing species has clearly increased in L. Võrtsjärv but much less in L. Peipsi. The increase of cyanobacterial dominance and the occurrence of water blooms can be caused by reduced N loading and decreased N/P ratio. The ecosystem of L. Võrtsjärv is very sensitive to water level fluctuations, which follow the North Atlantic Oscillation index refelecting climate changes in the northern hemisphere. In deeper L. Peipsi the direct influence of the water level is not so obvious.
| 1. Introduction | up | previous | next | last |
Regular investigation of two large shallow nonstratified lakes Peipsi ( 3 555 km 2 , mean depth 7.1m) and Võrtsjärv (270 km 2 , mean depth 2.8 m) started in the 1960s and up to now about 35-year time series data on water chemistry and biology have been collected. L. Peipsi is one of the most important lakes in Europe having the fourth largest surface area after Ladoga, Onega, and Vänern. Being located on the Estonian-Russian border, L. Peipsi is a transboundary waterbody and the largest international lake in Europe. L. Võrtsjärv is the largest lake belonging entirely to Estonia (Fig. 1).
Watershed of Lake Võrtsjärv (3 104 km 2 ) belongs to the basin of L. Peipsi (47 800 km 2 ). Riverine transport is the most important pathway for the input of nutrients into both lakes. For L. Võrtsjärv four main inflows, the rivers Väike Emajõgi, Õhne, Tarvastu and Tänassilma form 7075% of the water discharge, and 8085% of the total load of substances. In case of L. Peipsi, t he majority of phosphorous and nitrogen compounds (> 80%) is carried into the lake by the rivers Velikaya and Emajõgi (outflow from L. Võrtsjärv). The first carrying biologically treated sewage from the Russian town Pskov (201 400 inhabitants), the latter transporting wastewater from the Estonian town Tartu (101 200 inhabitants). The wastewater of Tartu remained untreated for a long time; the treatment plant has been in operation since the end of 1998. The contribution of the R. Emajõgi in the loading of total nitrogen (TN) and phosphorus (TP) into L. Peipsi from Estonian territory is ca. 70% (Fig. 2) . The Russian R. Velikaya is contributing about 65% of all nutrient loading into L. Peipsi (St å lnacke, 2001) and about 85% of that coming from the Russian territory.
Lake Peipsi consists of three parts. The area of the largest northern part, L. Peipsi sensu stricto , is 2,611 km 2 , mean depth 8.4 m and the maximum depth 12.9 m. The southern part, L. Pihkva with an area of 708 km 2 and mean depth of 3.8 m is connected with L. Peipsi by the narrow river‑shaped L. Lämmijärv having an area of 236 km 2 and mean depth of 2.6 m. The catchment area (47,800 km 2 ) involves parts of Estonian, Russian and Latvian territories. The volume of water in L. Peipsi is 25 km 3 at the long-term mean water level (30.00 m above sea level) and the mean residence time of water is about 2 years. The outflow, the River Narva, runs its waters into the Gulf of Finland.
L. Peipsi is an unstratified eutrophic lake, L. Lämmijärv has some dyseutrophic features, while L. Pihkva is a eutrophic, or even hypertrophic water body (Tab. 1).
The measurements of TP and TN in L. Peipsi started in 1985 and in the Estonian rivers in 1984. In Russian rivers and waters of L. Peipsi mainly the mineral forms of nutrients (ammonium, nitrites, nitrates, phosphates) have been tested. The data series on mineral N and P compounds in L. Peipsi started in 1968 and in the rivers in 1976. Regular biological data series date back to 1962.
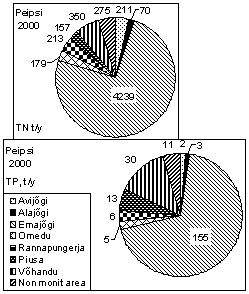
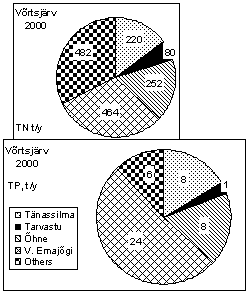
Figure 2. Nitrogen and phosphorus loading from Estonian watershed into L. Peipsi and Võrtsjärv in 2000, and the share of different rivers
Lake Võrtsjärv is highly eutrophic (Table. 1) receiving nutrients from four main rivers (Fig. 2). The outflow of L. Võrtsjärv, the River Emajõgi, has a small slope, only 3.8 cm km -1 . Opposite flow in the upper course of that river occurs almost every spring, while during about a half month period the lake has no outflow (Järvet, 1995).
Table 1. Indices reflecting trophic status of lakes Peipsi and Võrtsjärv (modified from Haberman et al., 1998; Laugaste et al., 2001; Starast et al., 2001)
Parameter |
Units |
L. Peipsi mean |
L. Pihkva |
L.Lämmi-järv |
L. Peipsi s.s. |
L. Võrtsjärv |
TP |
mg P m -3 |
42 |
63 |
53 |
35 |
54 |
TN |
mg N m -3 |
768 |
1010 |
923 |
678 |
1600 |
Chlorophyll a |
mg m -3 |
18 |
26 |
25 |
14 |
24 |
Secchi depth |
m |
1.8 |
1.3 |
1.4 |
1.8 |
1 |
A long-term sinusoidal fluctuation of the water level (WL) with a period of about 30 years is characteristic of both lakes (Fig. 3). A rather smooth continuous decrease (1928-1940) or increase (1940-1957; 1965-1990) within the period resembles a long-term trend and can be distinguished from it only in the context of a longer time series. Apparent periodicity is probably associated with large-scale fluctuations in solar activity and atmospheric processes, since it appears in the same way in large geographic regions. Similar periodicities with a spectral density maximum of 28-32 years have been found in the WL dynamics of the lakes of Saimaa, Ilmen and Onega (Masanova and Filatova, 1985), as well as for different hydrological elements of Lake Ladoga (20-30 years) (Malinina et al ., 1985) and Lake Müggelsee (Behrendt and Stellmacher, 1987). In addition to visual analysis of water level series, occurrence of hidden periodicity was studied with the help of the standard method of spectral analysis. Using Fourier ' s transformation of the correlation function, spectral density can be determined (Järvet, 2000). Altogether three groups of periodicity were detected: 1) short periods with a spectral peak at 3.6 years; 2) medium periods at 6.4 years; 3) long periods with a length between 24 and 32 years. The spectral peak at 25.6 years seems to have an exceptional position.
Shifts of the spectral behaviour of water level depends on the amount of precipitation. The corrected time-series of spatial mean annual precipitation in Estonia indicates a clear periodicity: cycles of 5060, 2533 and 57 years were detected (Jaagus, 1998). Temporal variability of evaporation is much lower than that of the runoff of rivers and water level of lakes. When annual precipitation in Estonia is less than 650 mm, then water level will depend on precipitation, and evaporation is stable (Järvet, 1998). When precipitation is higher, evaporation will increase with rising amount of precipitation. Based on the comparison of water balance elements, it can be concluded that the temporal variability of the water level of large lakes is much higher than the variability of precipitation.
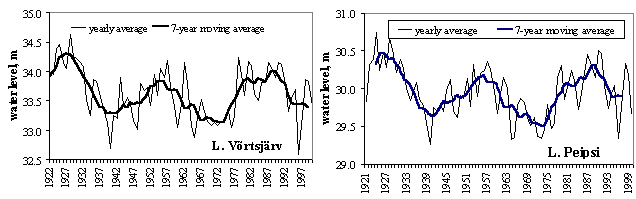
Figure 3. Long-term dynamics of the water level of lakes Võrtsjärv and Peipsi
In present paper we address the questions, how the runo ff, nutrient concentrations and riverine loadings have been changed during the last four decades, and how these changes have influenced the most sensitive part of the lake ecosystem phytoplankton. The consequences of global climate change are discussed a s well.
| 2. Changes in Riverline Discharges | up | previous | next | last |
Significant changes in the hydrological regime of rivers have taken place due to global climate changes and also changes in land-use and agricultural practice. Statistical analysis indicated that annual runoff remained almost changeless, however, the maximum and minimum discharges changed significantly. During the last 80 years, the amplitude of seasonal fluctuation of the runoff of the R. Emajõgi decreased - the shape of hydrographs became more flat (Fig. 4). The values of maximum discharges have decreased by 0.05 - 0.7 m 3 /s per year for most rivers, however, the trend was different for different years. Analysis showed that the range and values of runoff were bigger in the beginning of the 20 th century and more levelling after the 1960s. The decrease was most pronounced during the snowmelt period. It can be explained by shortening of the snow cover period and by the fact that frozen periods in winter often changed with thaw periods. Moreover, the beginning of spring melt-time shifted to the earlier date and the break-up of ice-cover occurs now on the average 15 days earlier than in three last centuries.
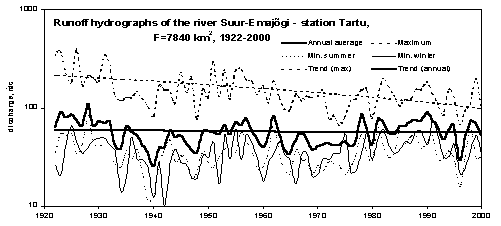
Figure 4. Long-term changes of the R. Emajõgi runoff
| 3. Changes of the River Water Chemistry | up | previous | next | last |
The present level of biological oxygen demand (BOD 7 ) in most rivers of L. Peipsi basin is quite low if compared with the 1980s ( Loigu et al., 2001) when the highest amount of wastewater was discharged into the rivers. The biochemical oxygen demand of natural river water, which is not directly influenced by human activity is, as a rule, less than 3.0 mgO 2 /l. BOD 7 3-5 mgO 2 /l indicates moderate human impact and values generally more than 5 mgO 2 /l indicate obvious pollution. BOD 7 level in rivers of L. Peipsi basin is generally classified as good (Class II) or moderate (Class III) during the last decade (Fig. 5). The fluctuation of annual 90% values is also quite low, which shows high efficiency of wastewater management in towns and villages in the basin.
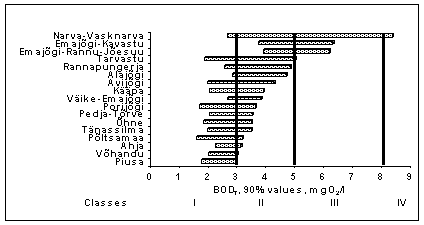
The rather high BOD 7 in the R. Emajõgi, especially at the outlet of L. Võrtsjärv (Rannu-Jõesuu station) is caused by high content of organic matter in L. Võrtsjärv. BOD 7 at Rannu-Jõesuu station is the highest in summer during the period of intensive photosyntesis due to the high amount of phytoplankton. Rivers discharging into L. Võrtsjärv are rather nutrient-rich. Organic matter produced in this highly eutrophic lake is the main factor causing relatively high biochemical oxygen demand together with high organic nitrogen and phosphorus concentrations in the R. Emajõgi. Therefore, the high level of BOD 7 at Rannu-Jõesuu station does not indicate direct organic pollution. The content of inorganic nitrogen and phosphorus at the outlet from L. Võrtsjärv is low because it is transferred to organic nitrogen and phosphorus during the photosyntesis process in the lake. As a result of phytoplankton degradation, upstream biochemical oxygen demand in summer and autumn (5-7 mgO 2 /l) can be even higher than downstream at Kavastu station, near the Tartu wastewater outlet.
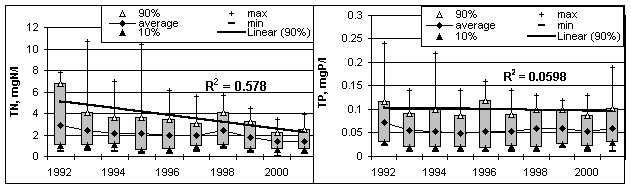
One of the main issues of Estonian water protection is the eutrophication of surface waters caused by the increased load of nutrients of anthropogenic origin. The eutrophication level of lakes depends on the nutrient content in rivers. The critical content of total nitrogen above which the eutrophication of rivers starts is 2.5 mgN l -1 (Loigu 1992). In Estonian rivers considered as background rivers, TN concentration was lower than the critical value in 98% of all samples, the mean concentration being 1.5 mgN l -1 . The nitrogen level in Estonian rivers is still quite high although the content of nitrogen has decreased in most rivers in recent years due to decreased agricultural pollution load. Concentration of nitrates was the highest in 1985-1987, when the use of fertilisers was very intensive and the major part of them leached into surface waters (Loigu & Leisk, 1996). The level of agricultural production and also nitrate concentration was the highest in 1984-1988, while by 1995-1999, which was the period of low agricultural production, the nitrogen concentration in rivers decreased (Fig. 6).
At the beginning of the R. Emajõgi the percentage of organic nitrogen and phosphorus is high and the content of mineral forms is low as mineral nutrients are transformed to organic form in the process of photosyntesis taking place in L. Võrtsjärv (Loigu & Leisk 1989). Organic N and P form on the average 72% of TN and TP, the content of mineral forms is higher only in spring flood periods (Fig. 7).
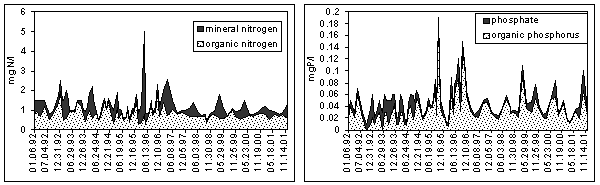
Fig. 7. Content of nitrogen and phosphorus at Rannu-Jõesuu station in the River Emajõgi in 1992-2001
| 4. Changes of Nutrient Loading | up | previous | next | last |
In the 1980s, the riverine discharge of nutrients into lakes Peipsi and Võrtsjärv increased while in the early 1990s a sharp decrease occurred, first of all in TN loadings (Fig. 8). This change was caused mainly by the collapse of the soviet-type agriculture characterised by heavy fertilization of fields often accompanied by substantial nutrient leakage into water bodies. A significant reduction in fertilizer use, typical for the transitional economy during recent years, diminished nutrient losses from the catchment. Compared to the level of the end of the 1980s, only 510% of N, P and K mineral fertilizers and 30% of the manure were applied in the agricultural lands at the end of the 1990s (Järvet et al .,2002). In addition to the change of land use pattern and fertilization intensity, variation in hydrological conditions plays an important role in nutrient and organic matter runoff from rural catchments. However, this influence is specific and varies for different nutrients. For instance, the annual water discharge correlated positively with the runoff of BOD 5 , NO 3 N and TIN (p<0.05), but there was no statistically significant correlation between stream discharge and NH 4 N and PO 4 P transport for monthly time series. As TN loading decreased faster than TP loading, the TN/TP ratio in the loadings decreased (Fig. 9).
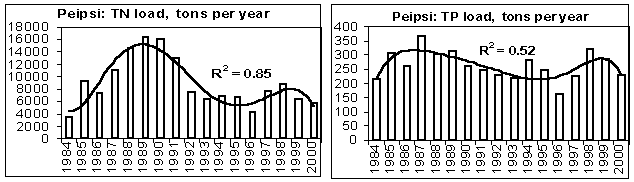
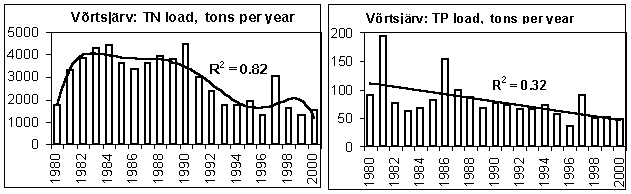
Figure 8. Annual runoff of total nitrogen (TN) and phosphorus (TP) into L. Peipsi from Estonian side in 19842000, and into L. Võrtsjärv in 19802000
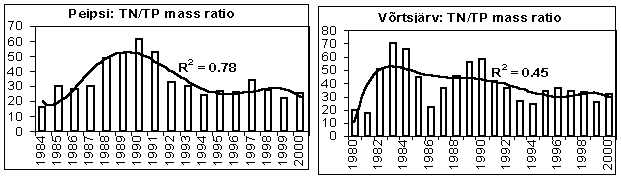
Figure 9. The ratio of total nitrogen and total phosphorus (TN/TP) in annual external loading of L. Peipsi (from Estonian part) and of L. Võrtsjärv
| 5. Consequences of the Changed Nutrient Loading on the Phytoplankton of Lakes | up | previous | next | last |
In L. Võrtsjärv average phytoplankton biomass in May-October 1963-2001 was 20 gWW m -3 and it has varied from 1 to 100 gWW m -3 ; in L. Peipsi s.s. the respective values in 1962-2001 were 10, 0.35 and 61 gWW m -3 . Many species of bloom-forming cyanobacteria are able to use molecular nitrogen at low N/P ratio if N is the limiting nutrient. Observing the changes in cyanobacterial dominance from the beginning of the 1960s in lakes Peipsi and Võrtsjärv, one can notice the increase in the late 1960s and early 1970s, decline in late 1970s and 1980s and a new increase in late 1990s in both lakes (Fig. 10). In L. Võrtsjärv changes are much less pronounced than in L. Peipsi. In L. Võrtsjärv dominant cyanobacterial species Limnothrix planktonica, L. redekei, Planktolyngbya limnetica are not able to fix N 2 , the main N 2 -fixing species, Aphanizomenon skujae , does not achieve dominating status . In L. Peipsi N 2 -fixing species Aphanizomenon flos-aquae and Gloeotrichia echinulata prevail in summer phytoplankton. Since the beginning of the 1990s, the biomass of N 2 fixing species has increased in both lakes (Fig. 11).
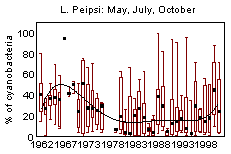
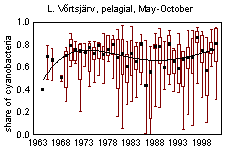
Figure 10. Changes in the dominance of cyanobacteria in phytoplankton biomass of L. Peipsi and L. Võrtsjärv since the 1960s.
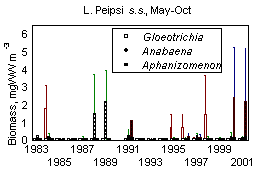
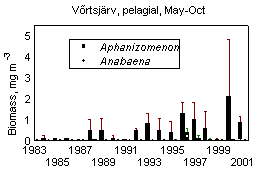
Figure 11. Biomass (median, quartiles, min, max) of N 2 -fixing cyanobacteria in lakes Peipsi and Võrtsjärv in 1983-2001
Cyanobacterial blooms are mainly attributed to the declining water quality. In lakes Peipsi and Võrtsjärv cyanobacteria constituted a substantial part of summer phytoplankton biomass already 40 years ago and their share declined in the conditions of extremely heavy nitrogen loading in the 1980s (Fig. 10). In both lakes cyanobacterial blooms have been documented since the beginning of the 20 th century (Mühlen & Schneider, 1920, Laugaste et al ., 2001). The increasing dominance of cyanobacteria and the occurrence of water blooms in both lakes can be caused by reduced nitrogen loading and decreased TN/TP ratio. Present situation is, however, closer to that of the 1960s when nitrogen load was lower and water blooms more common than in the 1980s at the time of extensive development of agriculture. So, water blooms may not always indicate eutrophication even if the blooms are heavy and bring about fish-kill as it happened during the bloom of G. echinulata in August, 2002.
| 6. Consequences of the Changes of Global Climate on Water Level and the State of the Ecosystem | up | previous | next | last |
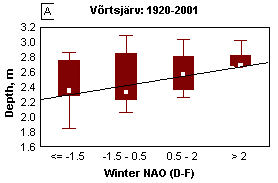
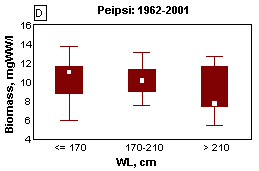
The North Atlantic Oscillation (NAO), which is defined through the variability of air pressure differences between the North (Iceland) and South (Azores), dictates climate variability over a large area of the Atlantic, North America and Europe, especially during winter (Hurrell et al ., 2001). Variation in heat and moisture transport between the Atlantic and surrounding continents affect water balance components of lakes, such as precipitations, riverine inflow and evaporation, resulting in changes of the water level. In both lakes Peipsi and Võrtsjärv mild winters associated with high North Atlantic Oscillation index (NAO) bring about higher water level (Fig. 11A&B). In very shallow L. Võrtsjärv where the annual amplitude of WL is 1.4 m and the absolute range is 3.2 m, the fluctuations of water level (WL) have strong influence on the ecosystem. In low-water years, better water column illumination and increased release of phosphorus from resuspended bottom sediments result in substantially higher phytoplankton biomass than in high-water years (Fig. 11C). In deeper L. Peipsi where seasonal and absolute WL variation ranges approximately
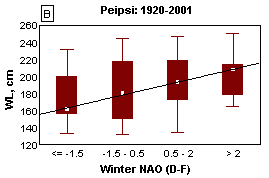
1/5 and 1/2 of the mean depth, respectively, the direct influence of the WL is not so obvious (Fig. 11D).
For the ecosystem of L. Võrtsjärv warmer and wetter climate could bring about higher water levels. The deeper the mixed water column, the lower is the average light intensity causing reduced phytoplankton biomass (Nõges & Nõges, 1998). In the deeper water both resuspension and denitrification rates are lower, the first reducing the phosphorus release from the bottom sediments and causing lower P concentration while the second reasoning the increased nitrogen concentration (Nõges & Nõges, 1999). Consequently, in warmer world the N/P ratio in Lake Võrtsjärv would be higher and N 2 -fixing cyanobacteria have less chances to develop (Fig. 12).
In the watersheds shorter freezing time of the soils and the increased amount of precipitation in warmer and wetter world can cause the higher nutrient leaching from the soils and accelerate the eutrophication of water bodies. We have noticed that in the 1990s, the spring peaks of the riverine nutrient discharges have been shifted to the earlier period (from the beginning April to mid March) than it used to be in the 1980s and earlier. Despite lower intensity of land use, the wintertime nutrient losses are almost as high as during former years because of increased water discharge and higher share of overland flow in winter.
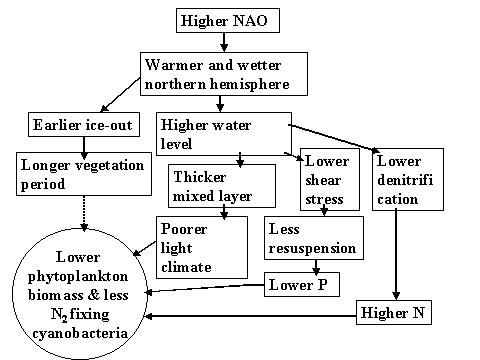
Figure 12. The assumed consequences of the global warming on the phytoplankton of Lake Võrtsjärv
| Acknowledgements | up | previous | next | last |
Funding for this research was provided by the project No 0370208s98 of Estonian Ministry of Education, by Estonian Science Foundation grant 4080, and by the European Union project MANTRA-East (contract EVK1-CT-2000-00076). We also used data obtained in frames of the State Monitoring Program of the Estonian Ministry of Environment.
| References | up | previous | next | last |
Behrendt, H. & Stellmacher, R. 1987. Long-term changes in water quality parameters of a shallow eutrophic lake and their relations to meteorologic and hydrologic elements. The Influence of Climate Change and Climatic Variability on the Hydrologic Regime and Water Resources (Proceedings of the Vancouver Symposium, August 1987). IAHS Publ. 168: 535-544.
Hurrell, J.W., Kushnir, Y. & Visbeck, M., 2001. The North Atlantic Oscillation. Science. 291: 603-605.
Jaagus, J. 1998. Climate fluctuations and trends in Estonia in the 20th century and possible climate change scenarios. In Climate Change Studies in Estonia (T. Kallaste and P. Kuldna, eds.), pp. 7-12. Stockholm Environment Institute Tallinn Centre.
Järvet, A., Mander, Ü., Kull, A., Kuusemets, V. Nutrient runoff change in a rural catchment in South Estonia. Large Rivers Vol. 13, No. 34. Arch. Hydrobiol. Suppl . 141/34, p. 305319, November 2002.
Järvet, A . Water regime of Lake Võrtsjärv. Estonia, Geographical Studies 8 (Eds. Tiia Kaare and Jaan-Mati Punning). Estonian Academy Publishers, Tallinn 2000, pp. 7288.
Järvet, A. 1998. Long-term changes in time series of water balance elements. In Country Case Study on Climate Change Impacts and Adaptation Assessments in the Republic of Estonia (A. Tarand and T. Kallaste, eds.), pp. 69-71. Stockholm Environment Institute Tallinn Centre.
Laugaste, R., Nõges, T., Nõges., P., Jastremskij, V.V., Milius, A., Ott, I., 2001. Algae. In: Pihu&Haberman (Eds.), Lake Peipsi. Flora and Fauna. Tartu: 31-49.
Malinina, T.I., Filatova, I.V. & Filatov, N.N. 1985. Long-term changes in the elements of water budget of Lake Ladoga. In Problemy issledovaniya krupnyh ozer. Leningrad: 79-81 (in Russian).
Masanova, M.D. & Filatova, I.V. 1985. Probability structure of interannual water level changes in North-Western lakes. In Problemy issledovaniya krupnyh ozer. Leningrad: 81-84 (in Russian).
Mühlen, M. von zur & Schneider, G. 1920. Der See Wirzjerw in Livland. Archiv für die Naturkunde des Ostbaltikums, 14: 1-156.
Nõges, P., Nõges, T. 1998. The effect of fluctuating water level on the ecosystem of Lake Võrtsjärv, Central Estonia. - Proc. Acad. Sci. Estonia. Ser. Ecology. 47. 2: 98-113.
Nõges, T. & P. Nõges. 1999. The effect of extreme water level decrease on hydrochemistry and phytoplankton in a shallow eutrophic lake. Hydrobiologia, 408/409: 277-283.
St å lnacke, P. (ed.) (2001) Nutrient Loads to Lake Peipsi. Environmental Monitoring of Lake Peipsi/ Chudskoe 1998- 99. Subproject Report Phase 2, Jordforsk Report No. 4/01, 65 pp.
| Address: | up | previous |
1 Institute of Zoology and Botany, Estonian Agricultural University, Võrtsjärv Limnological Station , 61101 Rannu, Tartumaa, Estonia. E-mail: tnoges@zbi.ee
2 Institute of Geography &
3 Institute of Zoology and Hydrobiology, University of Tartu, Vanemuise 46, 51014 Tartu, Estonia. E-mail: ajarvet@ut.ee
4 Tallinn Technical University, Department of Environmental Engineering, Ehitajate tee
5, 19086, Tallinn, Estonia. E -mail: ennloigu@edu.ttu.ee