|
THE BEHAVIOURAL
ECOLOGY OF SPECIES INTERACTIONS
|
| Back |
Plant-pollinator/seed-disperser, ant-plant,
plant-herbivore, and model-mimic interactions give rise to many evolutionary
(ultimate) and mechanistic (proximate) perspectives. The empirical studies on
these questions (both in the laboratory and in the field) and theoretical modeling
of the proximate mechanisms of herbivore foraging strategies that we are studying
in my lab are summarized in this issue.
Philosophy and Approaches
Hypotheses regarding expected strategies of each player in the interaction are
tested in our lab using the framework of the mechanisms involved in the interactions.
The proximate mechanisms are investigated using a multi - disciplinary approach,
using, for example, the tools of chemical analysis and mathematical and computer
modeling, apart from behavioral studies.
The plurality of approach is also mirrored in the diversity of the systems.
A brief summary of the major research projects is given in the next section.
|
Towards Understanding Foraging Movements in the Malabar
Giant Squirrel, Ratufa indica: Theoretical Modelling and Empirical Studies |
Introduction
The Malabar Giant Squirrel, Ratufa indica, is an endangered, almost completely
arboreal herbivore endemic to the closed canopy forests of India. Behavioural
ecology of this animal is being studied by conducting
extensive field studies, partly motivated by the conviction that a better understanding
of the biology of this animal will serve to guide conservation policies.
Empirical Studies
Previous work in the field has led to extensive information on the preferences
of the animal with respect to the biochemical compositions of its food resources.
Samples were collected, by means of continuous focal animal sampling, high -
resolution movement data of R. indica, along with behavioural observations
of resource utilization. This is complemented by spatio-temporal data on availability
of resources of differing quality. Empirically quantitating the spatial movement
constraints that the animal has to contend with while foraging is being done.
These constraints arise as a consequence of the animal being obligately arboreal,
which means that almost all the movements of the animal are within the canopy
of the forest, using the network of interconnected branches.
Theoretical Modelling
One of the primary motivations of the study is to elucidate the mechanisms by
which R.indica exercises its preferences for spatially distributed
resources. As in most studies involving endangered animals, spatial movement
data for R. indica is more readily available than information on
the behavioural mechanisms that guide the animal's movements. A methodology
for inferring behavioural mechanisms from movement data, therefore, would be
highly desirable. To this end, a model of animal movement was developed, which
takes into account behavioural mechanisms that guide an animal's movements while
predicting spatial movement patterns over time. Formerly, models used extensive
computer simulations to predict the expected patterns of movement. Compared
to this, now simple mathematical models are developed for greater reliability
and transparency. Thus, for example, the movement pattern of an animal using
an olfaction - guided navigation can be distinguished from that of an animal
using compass direction - based navigation. Using these models, one can, in
principle, infer some of the characteristics of the behavioural mechanisms from
spatio-temporal movement data. Conversely, given the known behavioural mechanisms
of movement, the probability of finding the animal at a point can be predicted.
Refinement of the models is in progress so as to incorporate more realistic
situations.
While these models are being developed with the objective of understanding R.
indica ' s foraging movements, they can be used to interpret spatial
movement data of any animal.
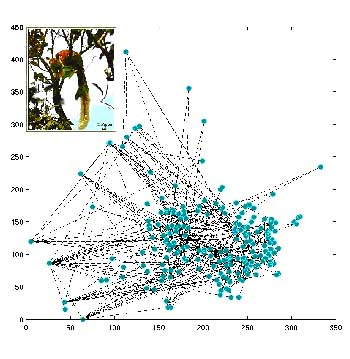
Theoretical representation of the territory of an individual
squirrel as a movement space graph (Figure 1), with the circles representing
the trees (nodes) and the lines (edges) representing the possibility of the
animal moving from one tree to another in a single step. Presence on each node
is considered as a 'state' of a Markov chain, with the transition matrix being
determined by the behavioural mechanisms and the spatial constraints. Scale
is in metres. Inset: The Malabar Giant Squirrel, Ratufa indica.
|
The Myrmecophyte Humboldtia brunonis (Fabaceae)
and its Invertebrate Associates
|
Myrmecophytes, which are plants that house ants, have an extraordinarily
diverse set of interactions with their ant partners. An interesting feature
of these interactions is the high frequency with which these interactions are
mutualistic, i.e., of benefit to both parties. Current focus is on the adaptive
strategies that are responsible for the evolution and maintenance of the mutualisms,
and their proximate manifestations. These questions in a system are being addressed,
which shows tremendous variation at different spatial scales both in the degree
of ant interaction as well as ant rewards provided by the plant, making it an
ideal system for asking evolutionary questions.
|
The system under investigation: Humboldtia brunonis
and its various associates
|
The association of H. brunonis with a wide range
of invertebrates is being studied and documented. Potential co-operations and
conflicts are hidden within these interactions churning out a system ideal for
investigation, rich in connections, exchanges and their respective consequences.
The central hypothesis currently being tested / investigated is that H. brunonis
invests a part of its resources in recruiting ant species that form a
line of defense against herbivory. From the proximate viewpoint, the resources
are in (at least) two forms:
Domatia, which are stem modifications that provide nesting space to invertebrates.
H. brunonis is polymorphic for the presence of domatia.
Extrafloral nectar, which is produced by nectaries on the sepals of buds
and young leaves.
Analyses of the chemical composition of the extrafloral nectar is done with
the objective of quantifying the reward provided to the ants. This is complimented
by behavioural studies on the ants to estimate the requirements and preferences
towards the individual components of the nectar.
The system, apart from showing considerable within site variation, also displays
between-site variation, with a gradation of increasing ant activity seen with
decreasing latitude. The causes of this variation form another subject of investigation.
Wherever it is found, H. brunonis also happens to be a dominant
understory tree. The hypothesis that this is largely due to vegetative propagation
is being tested using molecular studies.
 Domatia of Humboldtia brunonis |
 Domatia of Humboldtia brunonis |
 Extrafloral nectaries in Humboldtia brunonis |
 Ants on buds of H. brunonis |
Distribution of Humboldtia brunonis in the Western
Ghats
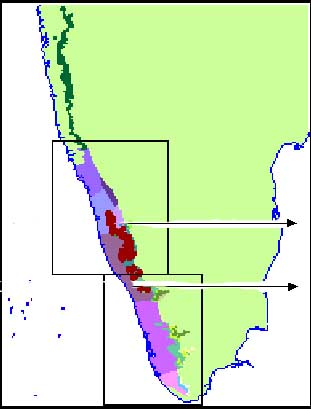 |
Wet Evergreen forests of the Western Ghats (low-elevation types,
abundant in H. brunonis)
Dipterocarpus indicus Humboldtia brunonis
|
| Behavioral Ecology of Ant-Mimicking Spiders |
Introduction
Myrmecomorphs are arthropods that have evolved a morphological
resemblance to ants. Myrmecomorphy in spiders is considered to be a type of
Batesian mimicry in which spiders gain protection from the predators through
morphological and behavioural similarity to aggressive or unpalatable ants.
Nine families of spiders have been recorded to exhibit mimicry, which include
Clubionidae, Salticidae, Thomisidae and Therididae. Myrmecomorphs are found
associated with a variety of terrestrial and arboreal ants of diverse habits
in terms of nesting preferences, trail forming habits and feeding ecology.
The System(s) under Investigation:
The mechanisms and the adaptive significance of mimicry in several species of ant-mimicking spiders are being investigated. The current focus is on the Myrmecomorphs of the family Salticidae, occurring in and around Bangalore. A number of diverse ant species have mimics that are closely related, though morphologically and behaviourally dissimilar. For example, the Weaver Ant, Oecophylla smaragdina has a mimic in the spider Myrmarachne plataleoides. The work also focuses on the intricate interactions between other ant species e.g. Camponotus and their spider mimics. The mimics are found in close proximity to their respective ant models.
Another interesting feature of myrmecophilic salticid spiders is their sexual dimorphism. The males have enlarged chelicerae which are also positioned parallel to the plane of the body unlike that of the female.
Questions Being Addressed
Given that the various morphs of spiders have evolved, from a common ancestor, under selection to mimic their respective models, the degree of morphological and behavioural similarity between each model - mimic pair is being investigated, as an estimate of adaptive change that a species can undergo under selection. Apart from behavioural studies, we are using the tools of digital image analysis towards this end. Along with predators, the spiders also need to avoid detection by the ant models. We are testing the hypothesis that the spiders achieve this by chemical mimicry, wherein the surface hydrocarbon profile, or the 'smell' of spiders is similar to that of their ant models. Chemical analysis (using mass spectrometry) is therefore being performed on these surface hydrocarbons, along with behavioural studies on the ants to estimate their ability to detect changes in these chemical 'signatures'. Field studies are being carried out to investigate the spatial proximity of each spider morph with its ant model.
Causes of sexual dimorphism in the system is another area of investigation. The dimorphism implies that the females are better mimics than the males. One of the interesting hypotheses we are pursuing is that the chelicerae of the males are objects of sexual selection, either as 'honest' signals of male quality by virtue of their being an impediment (the Handicap Principle - the 'good genes' hypothesis) or other, less adaptive reasons (the Fisherian 'good taste' hypothesis).
 The Weaver Ant, Oecophylla smaragdina (Model) |
 Male Spider Myrmarachne plataleoides (Mimic) |
 Female Spider Myrmarachne plataleoides (Mimic) |
| FIG AND FIG WASP MUTUALISM |
Introduction
Interaction between fig wasps and their host plants (Ficus,
Moraceae) range from mutualisms to parasitisms, and have served to test theories
of co-evolution, sex allocation, kin selection, speciation and conflicts of
resource management. All fig species are pollinated by mutualistic fig wasps
(Agaonidae, Agaoninae) that are characterized by extreme host specificity possessing
morphological adaptations and life cycles that depend entirely on host reproduction.
However this mutualism is also exploited by a large number of non-pollinating
wasps, which use figs as a breeding ground without offering pollination services.
Elucidating the ultimate and proximate mechanisms involved in the maintenance
of interactions between fig, fig -pollinator and parasite is one of the primary
research goals.
Fig -Pollinator- Parasite Interactions:
 Winged Female Wasp |
 Wingless Male Wasp |
 Non-Pollinating Wasp |
Resembling a fruit, the fig is an enclosed inflorescence (syconium) containing
many unisexual male and female flowers accessible to pollinators through an
opening called the ostiole. Pollen carrying females are attracted by host specific
fragrances released from receptive figs and lay eggs in a fraction of the flowers
they pollinate. Their offspring feed on fig endosperm as larvae and mate in
the fig cavity as adults. The inseminated female then leaves its natal fig with
the pollen in search of other receptive figs.
A diverse assemblage of nonpollinating wasps and parasitoids also co-exist.
Unlike the pollinators, fig-parasites oviposit within the flowers by piercing
from the outside of the syconium with long ovipositors, laying their eggs in
the ovaries but without the services of pollination, in turn competing with
the pollinators for space and food.
As part of the current research interests, now common Indian Ficus species (F.
racemosa, F. hispida, F. exasperata ) are being monitored
with the objective of characterizing the identity and behavior of their respective
pollinator and parasites.
Dioecy and Chemical Mimicry
In monoecious figs, all syconia in the tree are similar and produce both wasps
and seeds. However, in dioecious figs two separate trees occur - male and female
trees.
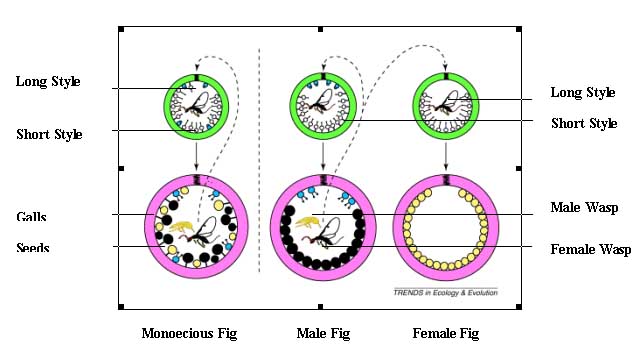
Male syconia contain male and short styled female flowers and produce many wasps,
but few if any seeds. By contrast, female syconia contain only long styled female
flowers and produce only seeds. While the entry of wasps into the female tree
is beneficial to the tree it acts as a burial ground for the wasps. This generates
a conflict of interest between the pollinator and the fig. Figs need agaonids
to enter both types of syconium, but agaonids can only produce offspring in
male syconia. From the perspective of the pollinator, therefore, the best strategy
would involve discriminating between male and female figs, and avoiding the
female figs altogether. From the fig's perspective, deceiving the pollinator
into believing that the female fig is actually male would be the best strategy.
The question, therefore, is whether there has been a selection on agaonids to
discriminate between male and female syconia and on figs to prevent this, and
what adaptations has each partner evolved so as to maximize its fitness.
The approach adopted to the above question is to investigate the mechanism by
which the pollinators recognize and locate figs. Previous studies have shown
figs release distinct chemical cues at different stages of their development,
which are recognized by its pollinating wasps. These chemical cues act as attractants
not only for the pollinators but also for frugivore animals, which help in dispersal
of seeds. If the volatiles released by the male and female figs differ, then
the wasps must be able to discriminate between them, while the figs should strive
to minimize these differences.
Volatiles collected from receptive male and female trees were subjected to GC-MS
analysis, so as to quantify differences in "chemical signatures". Behavioral
studies are being performed on the wasps using olfactometers to test their ability
to respond to differences in these signatures.
|
Dr. Renee M. Borges Centre for Ecological Sciences, Indian Institute of Science, Bangalore 560012, India Email: renee@ces.iisc.ac.in ,Url: http://ces.iisc.ac.in/hpg/renee/ Ph: 23600985 Extn: 105 |
TOP |