Reproduction |
Diatoms reproduce by two different modes, sexual and asexual.
Sexual reproduction:
The ability to reproduce sexually is closely associated with cell size of the diatom. It is the most prevalent method of restoring cell size, and it is often triggered when the size of a cell is less than a critical level (usually smaller than half of the original size for most species). The process of sexual reproduction is different for centric and pennate diatoms.
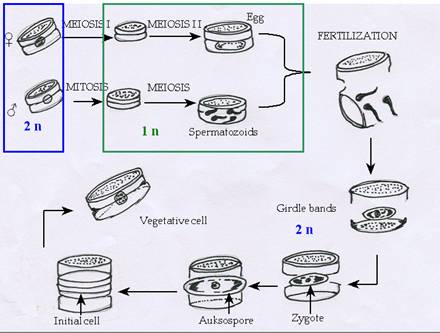
Sexual reproduction is oogamous in centric diatoms. A male gametangial cell first undergoes a series of differentiated divisions to form a limited number (usually four) of microspores, or sperm mother cells. The microspores then undergo meiosis to form flagellated sperm. Oogonial female cells produce one or two eggs; the resulting female protoplast facilitates sperm entry and favours the fertilisation. Fertilisation forms a zygote, which enlarges by water uptake to form an auxospore. The auxospore is often spherical in shape and with different valve morphology than normal valve. Division of the auxospore produces cells of the maximum size for that particular species.
Figure 8: Sexual reproduction in centric diatom
Sexual reproduction is isogamous in pennate diatoms. Gametangial cells pair up and undergo meiosis. Only one or two nuclei of the meiotic tetrad are sexually functional. The gametes are amoeboid and their emergence from their respective frustles is associated with the production of mucilage, which embeds the copulating cells. The gametes conjugate to produce usually one or two auxospores per pair of parents. As in centric diatoms, the auxospore divides to produce cells of restored size.
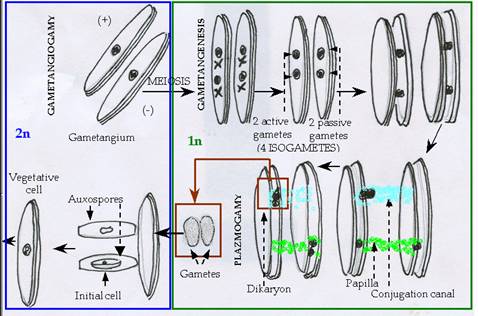
Figure 9: Sexual reproduction in pennate diatom
Asexual reproduction:
Diatoms have a unique "shrinking division" mode of asexual reproduction. The vegetative cell division involves a successive reduction in mean cell size as daughter cells are generated by the laying down of daughter thecae back and back within the parent cell, constrained by the relative rigidity of the cingulum wall. New sibling valves are usually formed in close juxtaposition to each other, so that their surface topographies may interact, e.g. where there is a depression on one sibling valve a raised area is formed on the other. Due to the rigidity of the test material, growth of the cell is impossible once the test is secreted. This has a number of consequences, one of these is that diatoms must be able to escape the inevitable size diminution if they are to survive. This is usually achieved via sexual reproduction in which gametes are formed and fuse outside the confines of the parent frustules, forming a zygote that expands to the species maximum, then laying down new silica valves to form new vegetative cells. Another consequence of size reduction, particularly for pennate diatoms, is that smaller cells may be mistaken for other taxa because valve outline and proportions may differ from that of larger cells. This size reduction process was independently reported by MacDonald (1869) and Pfitzer (1869) and has subsequently been known as MacDonald-Pfitzer rule. MacDonald-Pfitzer rule is not applicable to all the taxa (Jewson, 1992a, b), these size reduction size regeneration reproductive processes are responsible for controlling the size ranges of diatoms. In centric diatoms overall shape is unaltered, but in pennate diatoms reduction is greater along one axis so that shape as well as size is altered during mitosis.
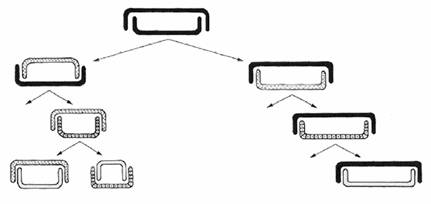
Figure 10: The MacDonaldPfitzer hypothesis (MacDonald 1869; Pfitzer 1869). At each division the new valve is always formed within the parental theca, causing the average size of the frustules in a population to slowly decrease.