Morphology |
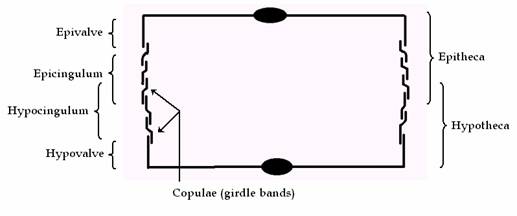
Diatom cells are contained within a unique silicate cell wall comprised of two separate valves. The biogenic silica that the cell wall is composed of is synthesised inside the diatom cell by the polymerisation (Polymerisation is a process of reacting monomer molecules together in a chemical reaction to form linear chains or a three-dimensional network of polymer chains) of silicic acid monomers. This material is then extruded to the cell exterior and added to the wall. Diatom cell walls are also called frustules, which are essentially bipartite structures, with an older and a younger half (valve); each with a series of linking bands (girdle bands or copulae), often comparable to a petri dish or soapbox. In the two halves of the cell, the hypotheca (the bottom portion in a soapbox) and the epitheca (the overlapping lid of the soapbox). The epitheca consists in turn of two parts, viz. a flat upper part with down-turned edges, called the epivalve (epivalva), and a ring or hoop like side wall, the epicingulum (upper girdle). Similarly, the hypotheca consists of a hypovalve (hypovalva) and a hypocingulum (lower girdle). The epicingulum and epivalve are separated by a suture, as are the hypocingulum and hypovalve (Figure 2 and 3).
Figure 2: Cross section through the frustule of a diatom, showing each half of the girdle (cingulum) that is composed of several girdle bands (copulae).
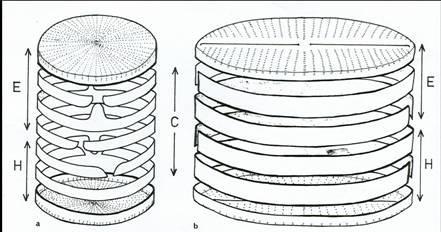
Figure 3: Diagram of diatom cells to show the relationships of the siliceous components (E = epitheca, epivalve + epicingulum; H = hypotheca, hypovalve + hypocingulum; C = cingulum, series of copulae) a, centric; b pennate. Each cell comprises two valves and a series of linking bands usually split rings. The band nearest the valve is often termed the valvocopoula. Copulae (=girdle bands) may have a tongue like extension (shown in a) which inserts into any space between the ends of the adjacent split copula.
The valves carry most of the taxonomic features used in standard flora and it is desirable in microscope preparations for the valve view to be clear to the observer. The cingulum comprised a single or sometimes multiple series of bands that are formed during the process of cell division, allowing the internal formation of daughter cells.
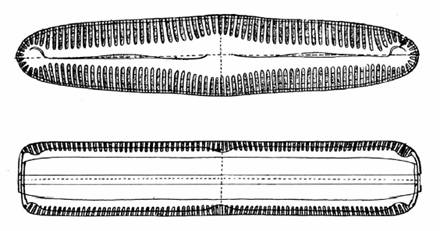
Diatoms may be divided into two major groups based on valve symmetry (Figure 5 and 6), those that are circular with essentially radial symmetry are the centric diatoms and those that are more elongate, with primarily bilateral symmetry are the pennate diatoms (Figure 5).
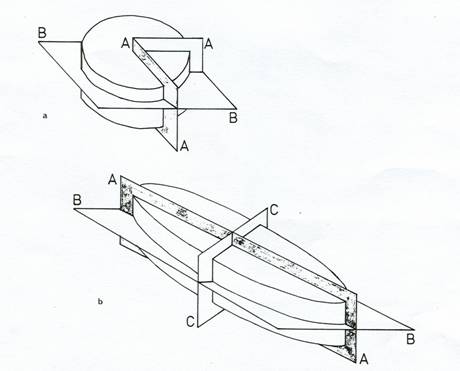
Figure 5: Diagrams to show axes and planes of symmetry used in diatom description
a) Centric diatom: AA = radial; BB = valvar. b) Pennate diatom: AA = apical; BB = valvar; CC = transapical.
The pennate diatoms are further sub-divided into two groups into the raphid and araphid diatoms, on the presence or absence of the raphe in the valve. The raphe is the one or two lateral slits alongside the sternum of the diatom cell. The raphe extends to cover a whole side of the cell initially, but during valve formation it may get altered. The alteration may split a single raphe into two raphes. A silica bridge, central nodule separates the double raphe.

Figure 6: Diagrams showing the general features of pennate diatoms (composite diagram) (Taylor et al, 2006)
The raphe may extend from pole to pole or may be incorporated into a different axis. Cross-linked bridges of silica termed fibulae support raphes. Some types of fibulae form canals beneath the raphe and it is termed as "canal raphes." Motility is the major function of the raphe and there are several ideas on the nature of diatom locomotion. One idea is that a polysaccharide is excreted from the raphe slits. All the motile diatoms possess a slit, the raphe through the wall and move only when the raphe is adjacent to a substratum to which the cell adheres. Mucilage secreted by the cell may be important in adhesion, and it has been proposed that the forces exerted by continuous secretion against the points of adhesion provide the motive force for locomotion (Drum & Hopkins, 1966, Harper & Harper, 1967, Harper, 1977). Another thought is that the ooze increases the flow of the lipid-coated underbody. The substratum attaches to the secretion by filaments of acid mucopolysaccharide and shortens like a primitive muscle tissue. It is presumed that there are ATPase and protein in the transmembrane structure.
Raphid diatoms are subdivided into monoraphid and biraphid taxa. Monoraphid diatoms have a functional raphe system on one valve only, the R valve. The other valve (P valve) has a solid raphe sternum since the raphe slit has been filled in with silica during morphogenesis. The position and path of the raphe slits and the presence or absence of associated ribs or struts are important features identification of the taxa. In some groups of diatoms, e.g. Nitzschia, Hantzschia, the raphe slits are positioned along one edge of the valve face, mostly raised along a ridge (keel) and spanned internally by struts (fibulae), which may appear as refractive lines or dots. In another group of diatoms, Surirellales, the fibulate raphe slits are positioned such that one runs along each long margin of the valve face, interrupted at both poles by nodules. Ontogenetically, one corresponds to the central nodule (the origin of the raphe system), the other to two fused polar nodules (Refer Figure 7 for different frustule features).
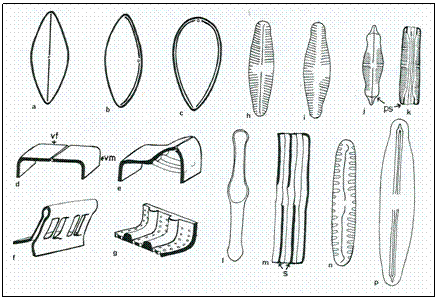
Figure 7: Diagrams to show frustule features. a-c, raphe paths with central nodules ( a : naviculoid along the centre of valve; b : nitzschiod displaced towards one valve margin; c : surirelloid raphe slits extending along each side of the valve. d-f 3D diagrams of raphe types ( d : naviculoid; e : nitzschoid with fibulae; f : surirelloid, raphe slit raised on a keel with additional supporting struts) g : diagram to show costaee; h : raphid; i : araphid; j : pseudosepta in valve and girdle ( k ); l : isolated girdle band of Tabellaria with septum; m : girdle view of half Tabellaria frustule showing the septa ( s ) projecting into the cell lumen; n : valve of Pinnularia borealis to show broad striae; p : valve of Frustulia vulgaris to show the internal ribs beside the raphe slits which fuse with the conspicuous helictoglossae to form porte-crayon endings.
In addition to being perforated by pores and/ or slits, some diatoms are ornamented with spines, often around the margin of the valve at the junction of the valve face and the valve mantle, or they may possess hollow processes (strutted or labiate processes) associated with the production of chitin fibres or mucilage fultoportula. These features may facilitate the formation of colonies. Specialised areas of simple pores (apical pore fields) often allow the secretion of mucilage pads or stalks for attachment to a substratum or to other cells in a colony. Some species have the ability to live as motile individuals or to attach to a substratum, depending on the circumstances.